

Genome sequences of four Ixodes species expands understanding of tick evolution
- PMID: 39838418
- PMCID: PMC11752866
- DOI: 10.1186/s12915-025-02121-1
Genome sequences of four Ixodes species expands understanding of tick evolution
Abstract
Background: Ticks, hematophagous Acari, pose a significant threat by transmitting various pathogens to their vertebrate hosts during feeding. Despite advances in tick genomics, high-quality genomes were lacking until recently, particularly in the genus Ixodes, which includes the main vectors of Lyme disease.
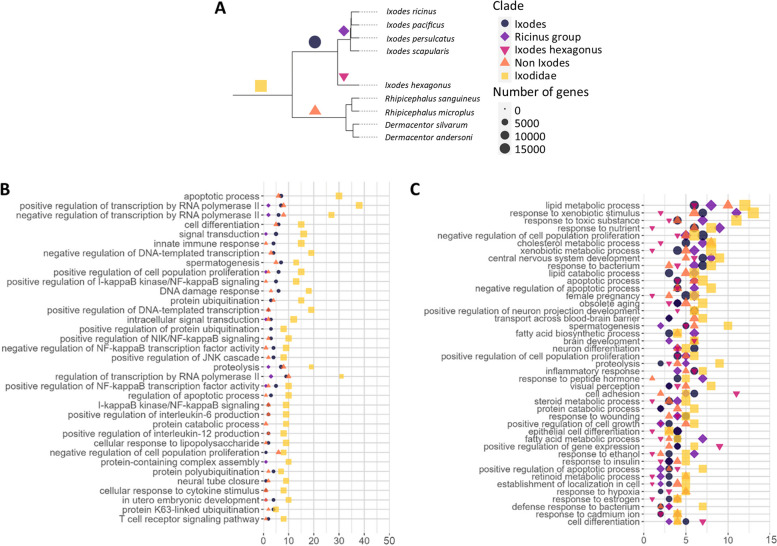
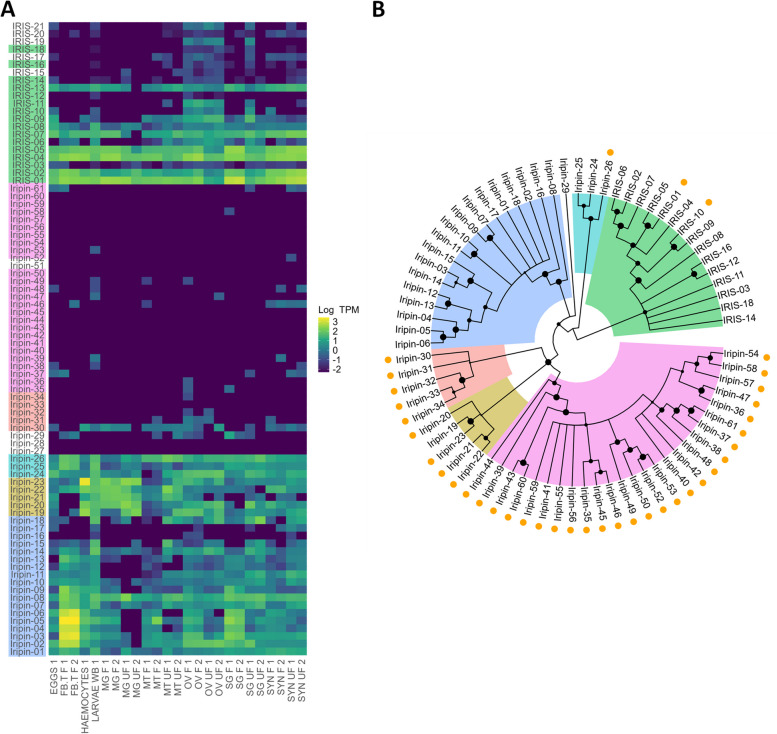
Results: Here, we present the genome sequences of four tick species, derived from a single female individual, with a particular focus on the European species Ixodes ricinus, achieving a chromosome-level assembly. Additionally, draft assemblies were generated for the three other Ixodes species, I. persulcatus, I. pacificus, and I. hexagonus. The quality of the four genomes and extensive annotation of several important gene families have allowed us to study the evolution of gene repertoires at the level of the genus Ixodes and of the tick group. We have determined gene families that have undergone major amplifications during the evolution of ticks, while an expression atlas obtained for I. ricinus reveals striking patterns of specialization both between and within gene families. Notably, several gene family amplifications are associated with a proliferation of single-exon genes-most strikingly for fatty acid elongases and sulfotransferases.
Conclusions: The integration of our data with existing genomes establishes a solid framework for the study of gene evolution, improving our understanding of tick biology. In addition, our work lays the foundations for applied research and innovative control targeting these organisms.
Keywords: Comparative genomics; Duplication; Hematophagy; Parasite; Retroposition.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The use of vertebrate animals (mice and rabbits) to maintain the I. ricinus colony at the University of Neuchâtel was performed following the Swiss legislation on animal experimentation. The commission that is part of the “Service de la Consommation et des Affaires Vétérinaires (SCAV)” of the canton of Vaud, Switzerland, evaluated and approved the ethics of this part of the study. The SCAV of the canton of Neuchâtel, Switzerland, issued the animal experimentation permit (NE05/2014). Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures





References
-
- Jongejan F, Uilenberg G. The global importance of ticks. Parasitology. 2004;129:S3-14. - PubMed
-
- Mans BJ, de Castro MH, Pienaar R, de Klerk D, Gaven P, Genu S, et al. Ancestral reconstruction of tick lineages. Ticks Tick-Borne Dis. 2016;7:509–35. - PubMed
-
- Dunlop JA. Geological history and phylogeny of Chelicerata. Arthropod Struct Dev. 2010;39:124–42. - PubMed
-
- Sharma PP, Kaluziak ST, Pérez-Porro AR, González VL, Hormiga G, Wheeler WC, et al. Phylogenomic interrogation of Arachnida reveals systemic conflicts in phylogenetic signal. Mol Biol Evol. 2014;31:2963–84. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
