

Uncovering a new family of conserved virulence factors that promote the production of host-damaging outer membrane vesicles in gram-negative bacteria
- PMID: 39840902
- PMCID: PMC11752146
- DOI: 10.1002/jev2.70032
Uncovering a new family of conserved virulence factors that promote the production of host-damaging outer membrane vesicles in gram-negative bacteria
Abstract
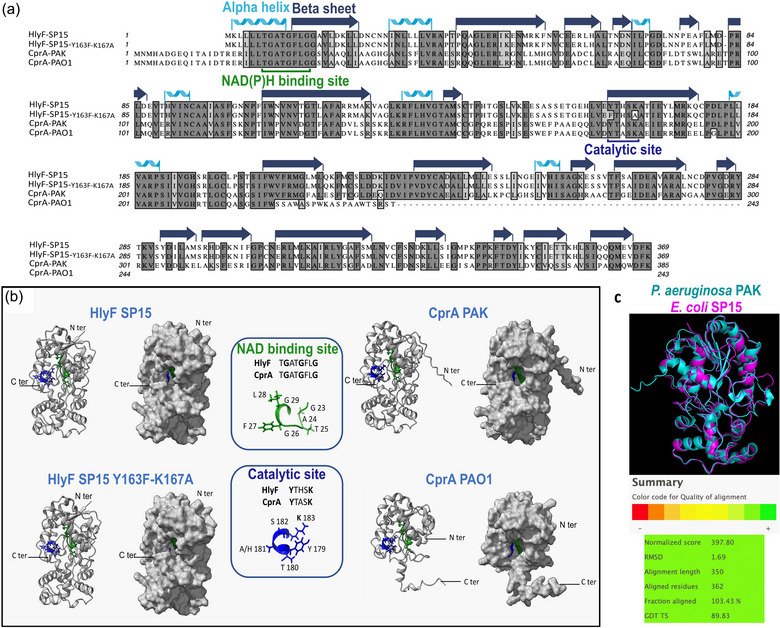
CprA is a short-chain dehydrogenase/reductase (SDR) that contributes to resistance against colistin and antimicrobial peptides. The cprA gene is conserved across Pseudomonas aeruginosa clades and its expression is directly regulated by the two-component system PmrAB. We have shown that cprA expression leads to the production of outer membrane vesicles (OMVs) that block autophagic flux and have a greater capacity to activate the non-canonical inflammasome pathway. In a murine model of sepsis, a P. aeruginosa strain deleted for cprA was less virulent than the wild-type (WT) strain. These results demonstrate the important role of CprA in the pathogenicity of P. aeruginosa. It is worth noting that CprA is also a functional ortholog of hemolysin F (HlyF), which is encoded by virulence plasmids of Escherichia coli. We have shown that other cryptic SDRs encoded by mammalian and plant pathogens, such as Yersinia pestis and Ralstonia solanacearum are functional orthologs of CprA and HlyF. These SDRs also induce the production of OMVs which block autophagic flux. This study uncovers a new family of virulence determinants in Gram-negative bacteria, offering potential for innovative therapeutic interventions and deeper insights into bacterial pathogenesis.
Keywords: OMVs; PmrAB; Pseudomonas aeruginosa; autophagy; inflammasome; pathogenicity; polymyxins.
© 2025 The Author(s). Journal of Extracellular Vesicles published by Wiley Periodicals LLC on behalf of International Society for Extracellular Vesicles.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- Bligh, E. G. , & Dyer, W. J. (1959). A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology, 37(8), 911–917. - PubMed
-
- Bricio‐Moreno, L. , Sheridan, V. H. , Goodhead, I. , Armstrong, S. , Wong, J. K. L. , Waters, E. M. , Sarsby, J. , Panagiotou, S. , Dunn, J. , Chakraborty, A. , Fang, Y. , Griswold, K. E. , Winstanley, C. , Fothergill, J. L. , Kadioglu, A. , & Neill, D. R. (2018). Evolutionary trade‐offs associated with loss of PmrB function in host‐adapted Pseudomonas aeruginosa. Nature Communications, 9, 2635. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
