

Attractor Landscape Analysis Reveals a Reversion Switch in the Transition of Colorectal Tumorigenesis
- PMID: 39840939
- PMCID: PMC11848608
- DOI: 10.1002/advs.202412503
Attractor Landscape Analysis Reveals a Reversion Switch in the Transition of Colorectal Tumorigenesis
Abstract
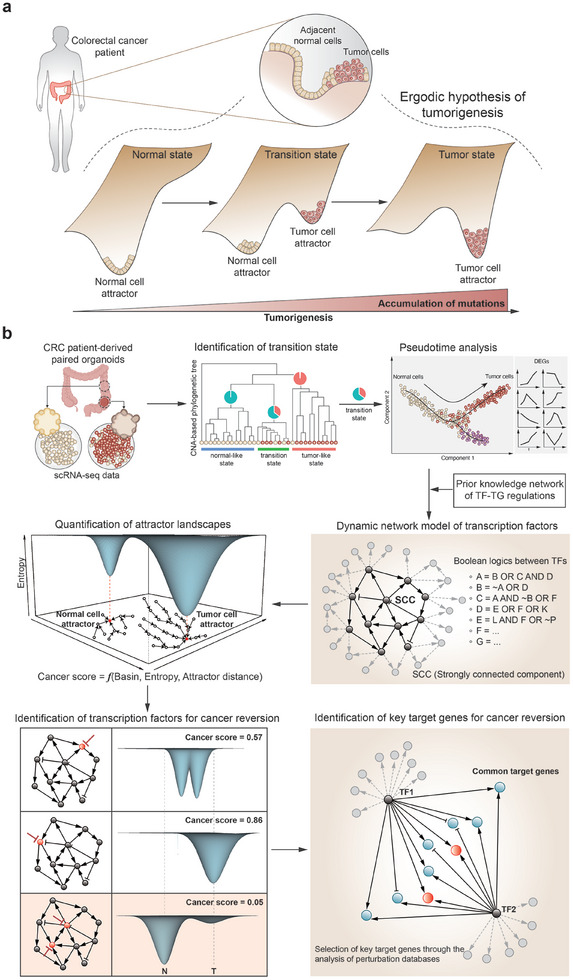
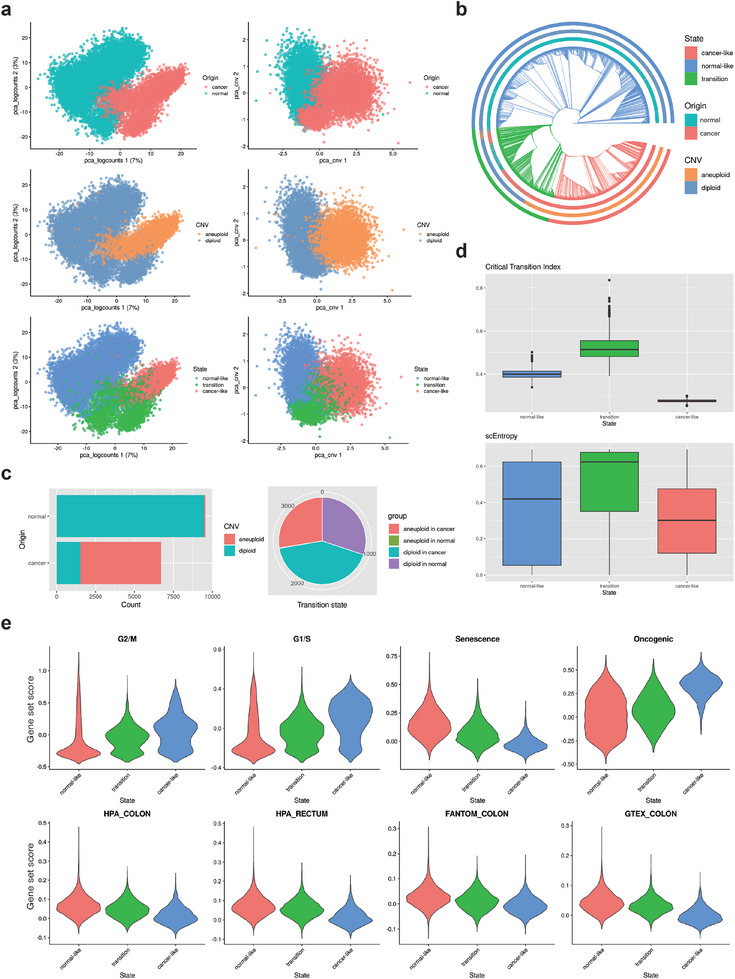
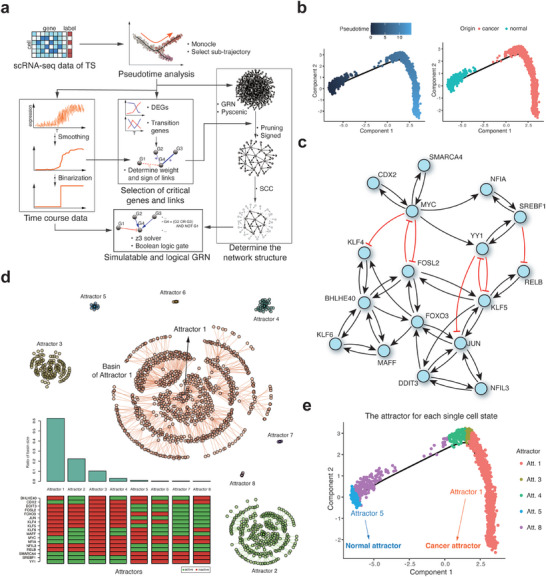
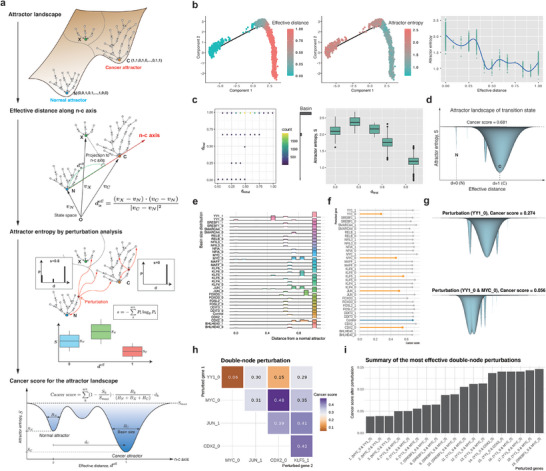
A cell fate change such as tumorigenesis incurs critical transition. It remains a longstanding challenge whether the underlying mechanism can be unraveled and a molecular switch that can reverse such transition is found. Here a systems framework, REVERT, is presented with which can reconstruct the core molecular regulatory network model and a reversion switch based on single-cell transcriptome data over the transition process is identified. The usefulness of REVERT is demonstrated by applying it to single-cell transcriptome of patient-derived matched organoids of colon cancer and normal colon. REVERT is a generic framework that can be applied to investigate various cell fate transition phenomena.
Keywords: attractor landscape analysis; cancer reversion; colon cancer; critical transition; dynamic network model; patient‐derived organoid; single cell transcriptome data.
© 2025 The Author(s). Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
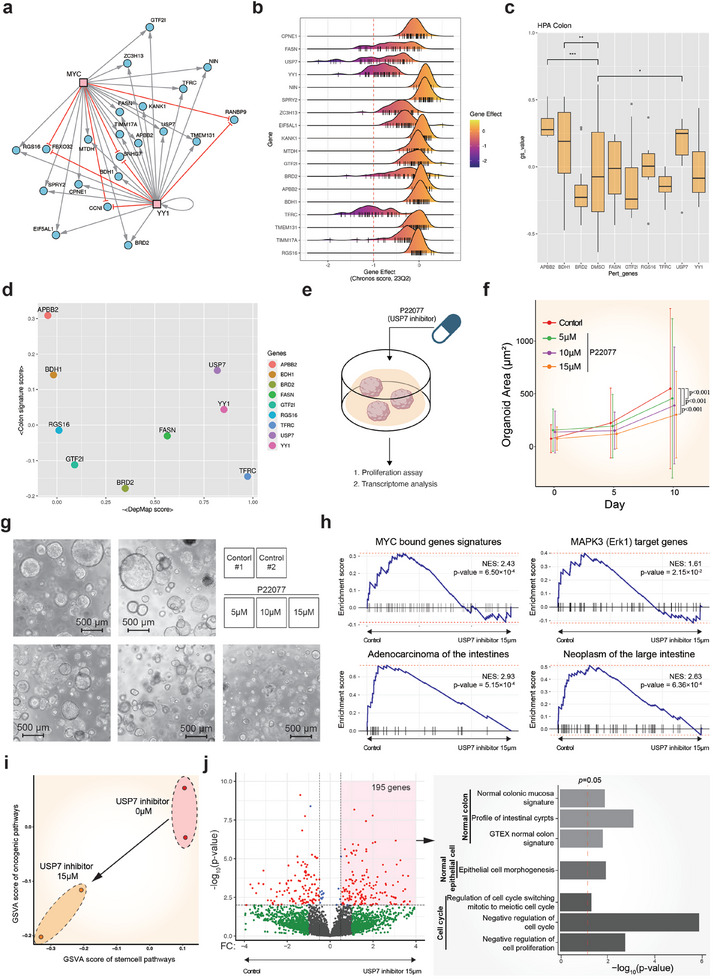
References
-
- a) Mojtahedi M., Skupin A., Zhou J., Castano I. G., Leong‐Quong R. Y., Chang H., Trachana K., Giuliani A., Huang S., PLoS Biol. 2016, 14, e2000640; - PMC - PubMed
- b) Poovathingal S. K., Kravchenko‐Balasha N., Shin Y. S., Levine R. D., Heath J. R., Small 2016, 12, 1425; - PMC - PubMed
- c) Zhong J., Han C., Chen P., Liu R., Brief Bioinform. 2023, 24, bbad366; - PubMed
- d) Gan Y., Guo C., Guo W., Xu G., Zou G., Brief Bioinform. 2022, 23, bbac225; - PubMed
- e) Sha Y., Wang S., Zhou P., Nie Q., Nucleic Acids Res. 2020, 48, 9505; - PMC - PubMed
- f) Giuliani A., Tsuchiya M., Yoshikawa K., Entropy 2017, 20, 13. - PMC - PubMed
-
- Askanazy M., Verhandlungen der Deutschen Pathologischen Gesellschaft 1907, 11, 44.
-
- a) Telerman A., Amson R., Nat. Rev. Cancer 2009, 9, 206; - PubMed
- b) Lo‐Coco F., Avvisati G., Vignetti M., Thiede C., Orlando S. M., Iacobelli S., Ferrara F., Fazi P., Cicconi L., Di Bona E., Specchia G., Sica S., Divona M., Levis A., Fiedler W., Cerqui E., Breccia M., Fioritoni G., Salih H. R., Cazzola M., Melillo L., Carella A. M., Brandts C. H., Morra E., von Lilienfeld‐Toal M., Hertenstein B., Wattad M., Lubbert M., Hanel M., Schmitz N., et al., d. A. Gruppo Italiano Malattie Ematologiche, G. German‐Austrian Acute Myeloid Leukemia Study, L. Study Alliance , N. Engl. J. Med. 2013, 369, 111; - PubMed
- c) Dow L. E., O'Rourke K. P., Simon J., Tschaharganeh D. F., van Es J. H., Clevers H., Lowe S. W., Cell 2015, 161, 1539; - PMC - PubMed
- d) Lee S., Lee C., Hwang C. Y., Kim D., Han Y., Hong S. N., Kim S. H., Cho K. H., Mol. Cancer Res. 2020, 18, 118; - PubMed
- e) Shin D., Cho K. H., Exp. Mol. Med. 2023, 55, 692. - PMC - PubMed
-
- Choi S. R., Hwang C. Y., Lee J., Cho K. H., Cancer Res. 2022, 82, 320. - PubMed
MeSH terms
Grants and funding
- 2022R1A2C1010532/Korean Government, the Ministry of Science and ICT
- 2023R1A2C3002619/Korean Government, the Ministry of Science and ICT
- RS-2024-00405360/Korean Government, the Ministry of Science and ICT
- HI21C0071/Ministry of Health & Welfare, Republic of Korea
- NCC-2410750/National Cancer Center/Republic of Korea
LinkOut - more resources
Full Text Sources
Medical