

Itaconate mechanism of action and dissimilation in Mycobacterium tuberculosis
- PMID: 39841148
- PMCID: PMC11789021
- DOI: 10.1073/pnas.2423114122
Itaconate mechanism of action and dissimilation in Mycobacterium tuberculosis
Abstract
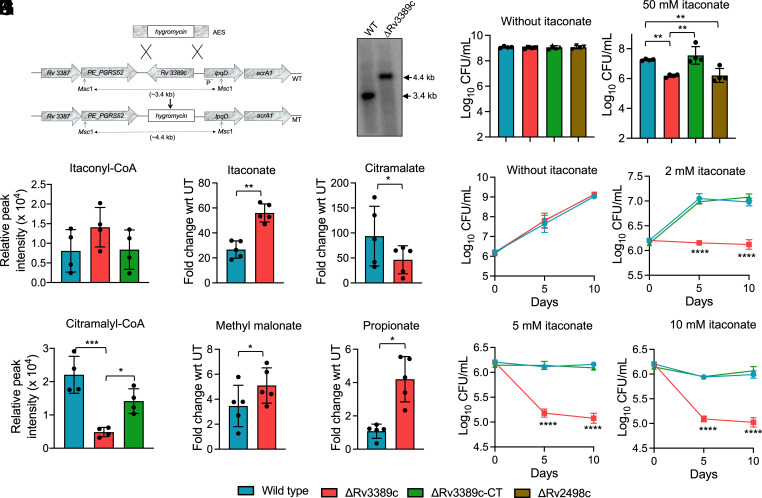
Itaconate, an abundant metabolite produced by macrophages upon interferon-γ stimulation, possesses both antibacterial and immunomodulatory properties. Despite its crucial role in immunity and antimicrobial control, its mechanism of action and dissimilation are poorly understood. Here, we demonstrate that infection of mice with Mycobacterium tuberculosis increases itaconate levels in lung tissues. We also show that exposure to itaconate inhibits M. tuberculosis growth in vitro, in macrophages, and mice. We report that exposure to sodium itaconate (ITA) interferes with the central carbon metabolism of M. tuberculosis. In addition to the inhibition of isocitrate lyase (ICL), we demonstrate that itaconate inhibits aldolase and inosine monophosphate (IMP) dehydrogenase in a concentration-dependent manner. Previous studies have shown that Rv2498c from M. tuberculosis is the bona fide (S)-citramalyl-CoA lyase, but the remaining components of the pathway remain elusive. Here, we report that Rv2503c and Rv3272 possess itaconate:succinyl-CoA transferase activity, and Rv2499c and Rv3389c possess itaconyl-CoA hydratase activity. Relative to the parental and complemented strains, the ΔRv3389c strain of M. tuberculosis was attenuated for growth in itaconate-containing medium, in macrophages, mice, and guinea pigs. The attenuated phenotype of ΔRv3389c strain of M. tuberculosis is associated with a defect in the itaconate dissimilation and propionyl-CoA detoxification pathway. This study thus reveals that multiple metabolic enzymes are targeted by itaconate in M. tuberculosis. Furthermore, we have assigned the two remaining enzymes responsible for the degradation of itaconic acid into pyruvate and acetyl-CoA. Finally, we also demonstrate the importance of enzymes involved in the itaconate dissimilation pathway for M. tuberculosis pathogenesis.
Keywords: Mycobacterium tuberculosis; dissimilation; itaconate; pathogenesis.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- Bagcchi S., WHO’s global tuberculosis report 2022. Lancet Microbe 4, e20 (2023). - PubMed
-
- Hubert C. B., de Carvalho L. P. S., Convergence and divergence in the metabolic network of Mycobacterium tuberculosis. Curr. Opin. Syst. Biol. 28, 100384 (2021).
-
- Munoz-Elias E. J., Upton A. M., Cherian J., McKinney J. D., Role of the methylcitrate cycle in Mycobacterium tuberculosis metabolism, intracellular growth, and virulence. Mol. Microbiol. 60, 1109–1122 (2006). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
