

Roles for the canonical polarity machinery in the de novo establishment of polarity in budding yeast spores
- PMID: 39841544
- PMCID: PMC11974964
- DOI: 10.1091/mbc.E24-07-0303
Roles for the canonical polarity machinery in the de novo establishment of polarity in budding yeast spores
Abstract
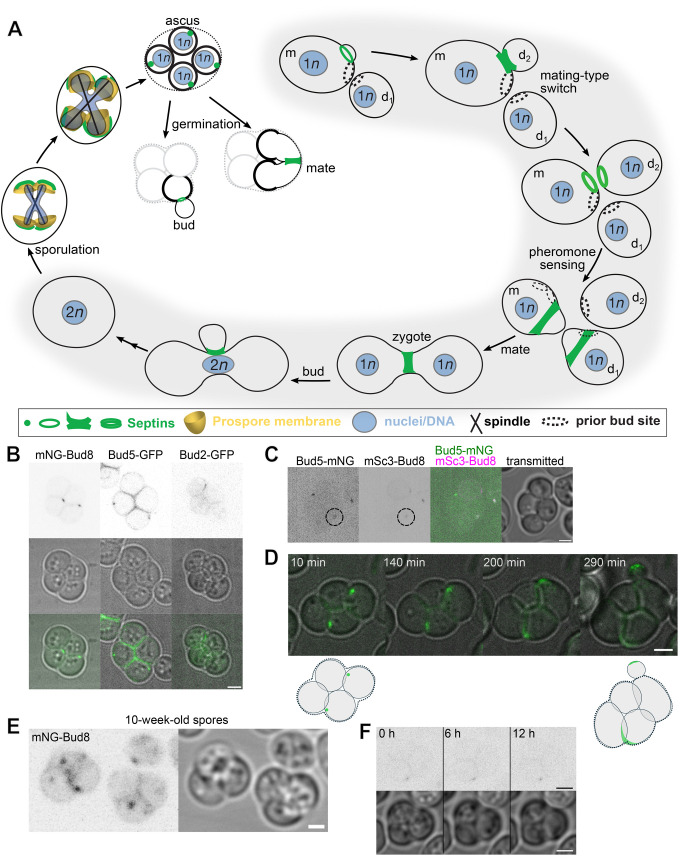
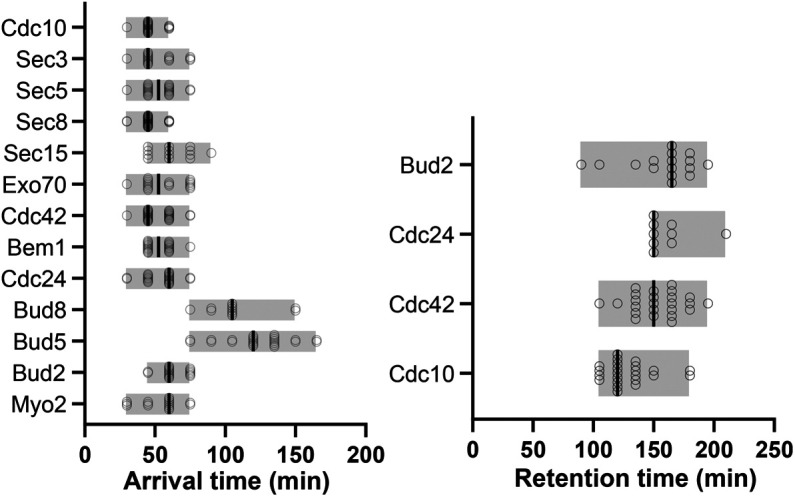
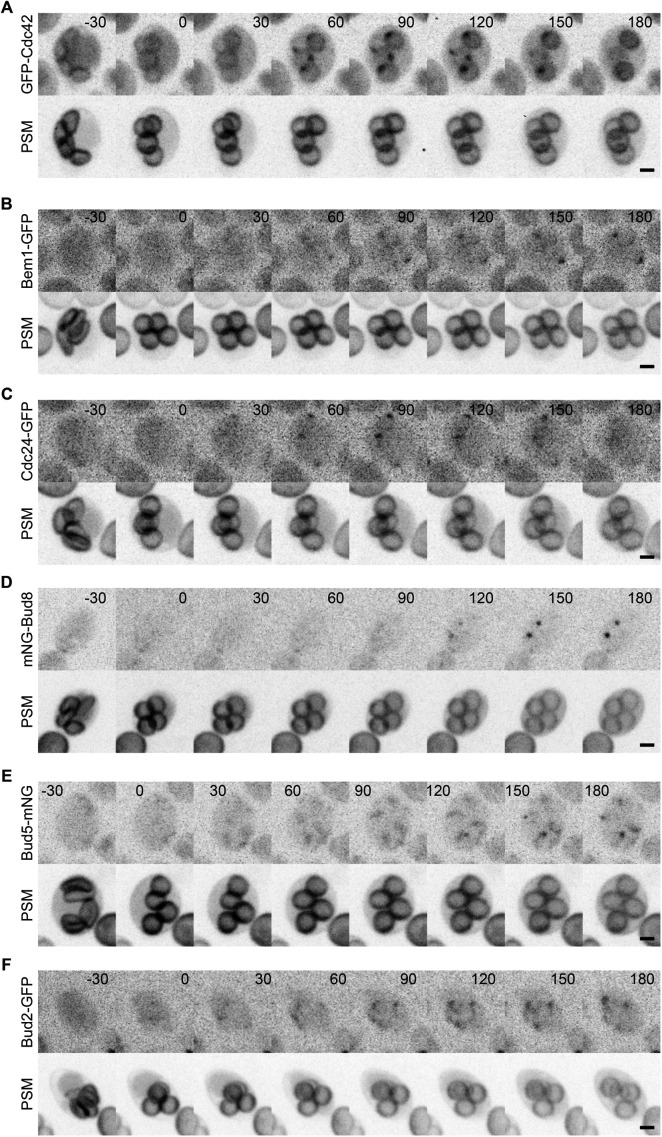
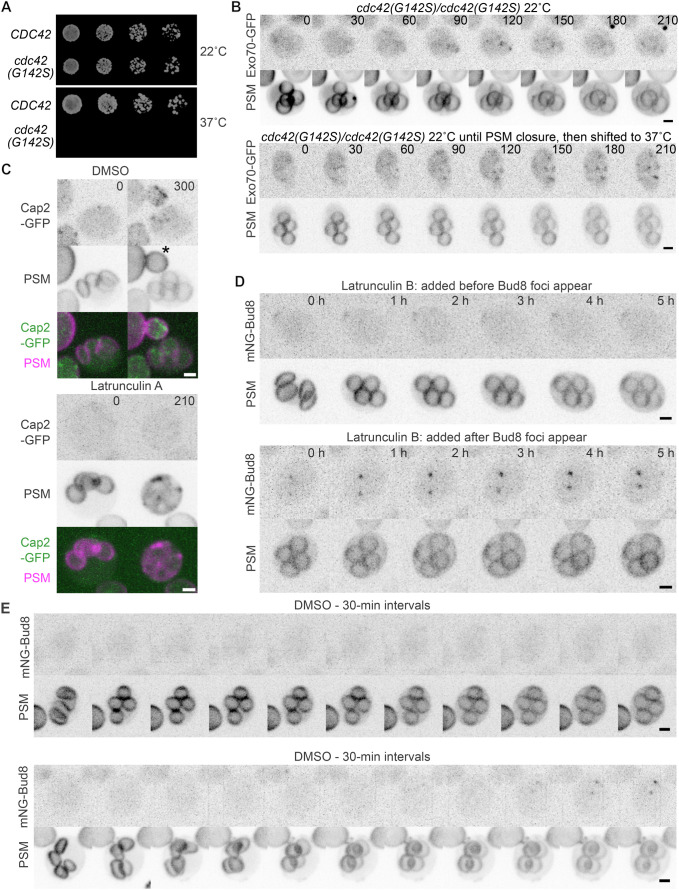
The yeast Saccharomyces cerevisiae buds at sites predetermined by cortical landmarks deposited during prior budding. During mating between haploid cells in the lab, external pheromone cues override the cortical landmarks to drive polarization and cell fusion. By contrast, in haploid gametes (called spores) produced by meiosis, a predetermined polarity site drives initial polarized morphogenesis independent of mating partner location. Spore membranes are made de novo so existing cortical landmarks were unknown, as were the mechanisms by which the spore polarity site is made and how it works. We find that the landmark canonically required for distal budding, Bud8, stably marks the spore polarity site along with Bud5, a GEF for the GTPase Rsr1 that canonically links cortical landmarks to the conserved Cdc42 polarity machinery. Cdc42 and other GTPase regulators arrive at the site during its biogenesis, after spore membrane closure but apparently at the site where membrane synthesis began, and then these factors leave, pointing to the presence of discrete phases of maturation. Filamentous actin may be required for initial establishment of the site, but thereafter Bud8 accumulates independent of actin filaments. These results suggest a distinct polarization mechanism that may provide insights into gamete polarization in other organisms.
Conflict of interest statement
Conflicts of interest: The authors declare no competing financial interests.
Figures




Update of
-
Roles for the canonical polarity machinery in the de novo establishment of polarity in budding yeast spores.bioRxiv [Preprint]. 2025 Jan 6:2024.08.29.610423. doi: 10.1101/2024.08.29.610423. bioRxiv. 2025. Update in: Mol Biol Cell. 2025 Mar 01;36(3):ar28. doi: 10.1091/mbc.E24-07-0303. PMID: 39257763 Free PMC article. Updated. Preprint.
References
-
- Amberg D, Burke D, Strathern J (2005). Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual, 2005 Edition, Cold Spring Harbor Laboratory Press.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
