

Motif distribution and DNA methylation underlie distinct Cdx2 binding during development and homeostasis
- PMID: 39843425
- PMCID: PMC11754732
- DOI: 10.1038/s41467-025-56187-0
Motif distribution and DNA methylation underlie distinct Cdx2 binding during development and homeostasis
Abstract
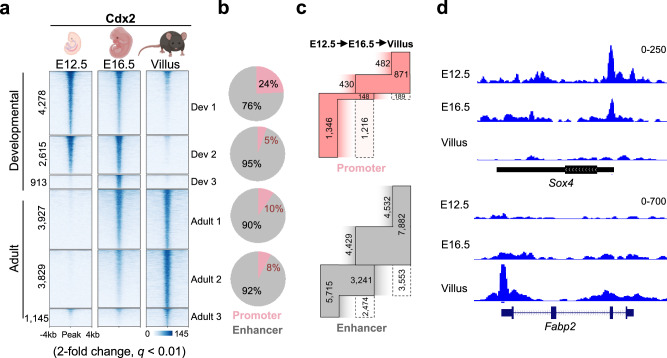
Transcription factors guide tissue development by binding to developmental stage-specific targets and establishing an appropriate enhancer landscape. In turn, DNA and chromatin modifications direct the genomic binding of transcription factors. However, how transcription factors navigate chromatin features to selectively bind a small subset of all the possible genomic target loci remains poorly understood. Here we show that Cdx2-a lineage defining transcription factor that binds distinct targets in developing versus adult intestinal epithelial cells-has a preferential affinity for a non-canonical CpG-containing motif in vivo. A higher frequency of this motif at embryonic Cdx2 targets and methylated state of the CpG during development enables selective Cdx2 binding and activation of developmental enhancers and genes. In adult cells, demethylation at these enhancers prevents ectopic Cdx2 binding, instead directing Cdx2 to its canonical motif without a CpG. This shift in Cdx2 binding facilitates Ctcf and Hnf4 recruitment, establishing super-enhancers during development and homeostatic enhancers in adult cells, respectively. Induced DNA methylation in adult mouse epithelium or cultured cells recruits Cdx2 to developmental targets, promoting corecruitment of partner transcription factors. Thus, Cdx2's differential CpG motif preferences enable it to navigate distinct DNA methylation profiles, activating genes specific to appropriate developmental stages.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures






Similar articles
-
DNA methylation-dependent and -independent binding of CDX2 directs activation of distinct developmental and homeostatic genes.bioRxiv [Preprint]. 2024 Mar 7:2024.02.11.579850. doi: 10.1101/2024.02.11.579850. bioRxiv. 2024. PMID: 38405700 Free PMC article. Preprint.
-
Intestinal master transcription factor CDX2 controls chromatin access for partner transcription factor binding.Mol Cell Biol. 2013 Jan;33(2):281-92. doi: 10.1128/MCB.01185-12. Epub 2012 Nov 5. Mol Cell Biol. 2013. PMID: 23129810 Free PMC article.
-
The lineage-specific transcription factor CDX2 navigates dynamic chromatin to control distinct stages of intestine development.Development. 2019 Mar 1;146(5):dev172189. doi: 10.1242/dev.172189. Development. 2019. PMID: 30745430 Free PMC article.
-
Transcription factors GATA4 and HNF4A control distinct aspects of intestinal homeostasis in conjunction with transcription factor CDX2.J Biol Chem. 2015 Jan 16;290(3):1850-60. doi: 10.1074/jbc.M114.620211. Epub 2014 Dec 8. J Biol Chem. 2015. PMID: 25488664 Free PMC article.
-
Extending the functions of the homeotic transcription factor Cdx2 in the digestive system through nontranscriptional activities.World J Gastroenterol. 2015 Feb 7;21(5):1436-43. doi: 10.3748/wjg.v21.i5.1436. World J Gastroenterol. 2015. PMID: 25663763 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
- K01 DK113067/DK/NIDDK NIH HHS/United States
- R03 DK134799/DK/NIDDK NIH HHS/United States
- T32 HD060549/HD/NICHD NIH HHS/United States
- K01DK113067, R03DK134799/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
