

A map of the rubisco biochemical landscape
- PMID: 39843747
- PMCID: PMC11839469
- DOI: 10.1038/s41586-024-08455-0
A map of the rubisco biochemical landscape
Erratum in
-
Author Correction: A map of the rubisco biochemical landscape.Nature. 2025 Feb;638(8052):E47. doi: 10.1038/s41586-025-08707-7. Nature. 2025. PMID: 39930266 Free PMC article. No abstract available.
Abstract
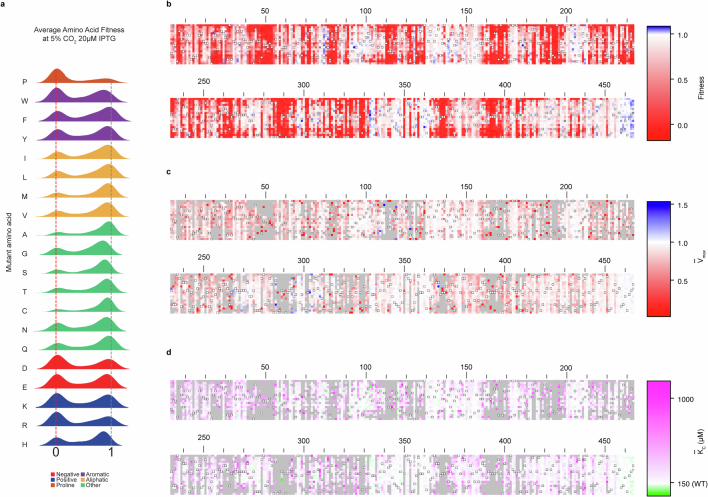
Rubisco is the primary CO2-fixing enzyme of the biosphere1, yet it has slow kinetics2. The roles of evolution and chemical mechanism in constraining its biochemical function remain debated3,4. Engineering efforts aimed at adjusting the biochemical parameters of rubisco have largely failed5, although recent results indicate that the functional potential of rubisco has a wider scope than previously known6. Here we developed a massively parallel assay, using an engineered Escherichia coli7 in which enzyme activity is coupled to growth, to systematically map the sequence-function landscape of rubisco. Composite assay of more than 99% of single-amino acid mutants versus CO2 concentration enabled inference of enzyme velocity and apparent CO2 affinity parameters for thousands of substitutions. This approach identified many highly conserved positions that tolerate mutation and rare mutations that improve CO2 affinity. These data indicate that non-trivial biochemical changes are readily accessible and that the functional distance between rubiscos from diverse organisms can be traversed, laying the groundwork for further enzyme engineering efforts.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: D.F.S. is a co-founder and scientific advisory board member of Scribe Therapeutics. The other authors declare no competing interests.
Figures
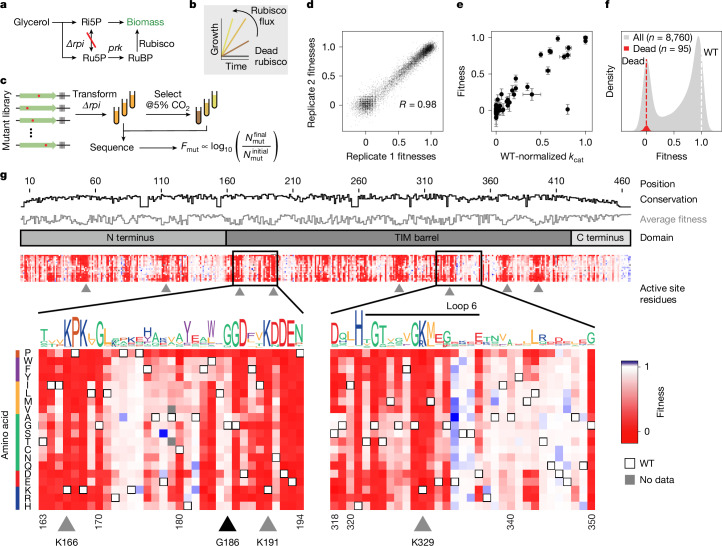
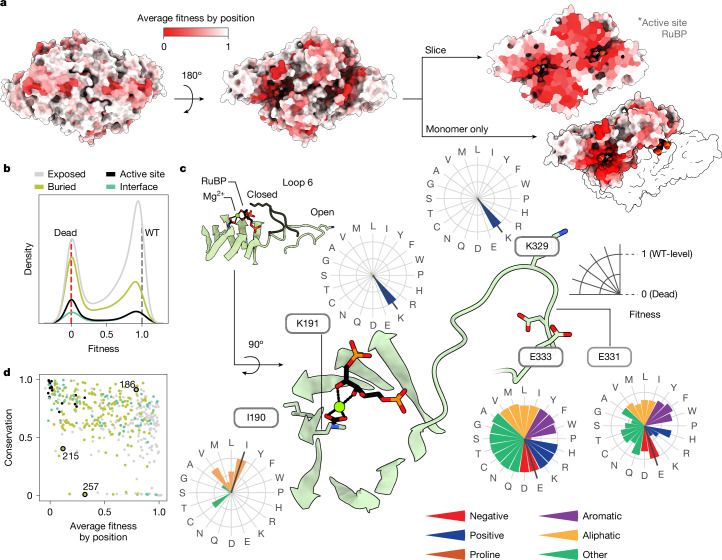
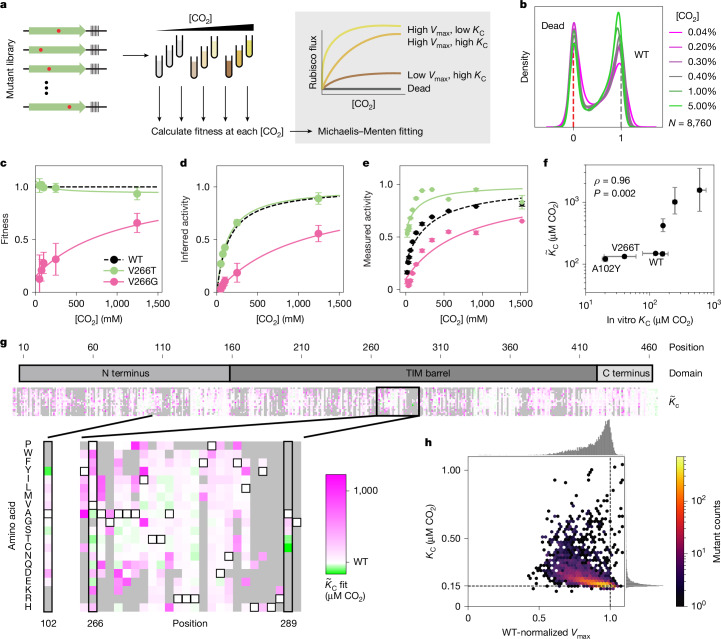
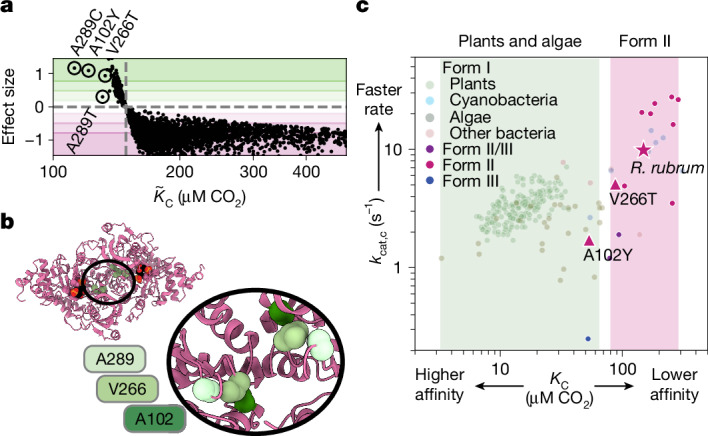

 . (PDB: 9RUB). Centre, Ribbon diagram of one monomer with the 3 subdomains labelled. View is of the interfacial side. Right, Close-up view of the active site. Closed form of loop 6 is from the 8RUC structure. Active site residues and RuBP substrate are labelled.
. (PDB: 9RUB). Centre, Ribbon diagram of one monomer with the 3 subdomains labelled. View is of the interfacial side. Right, Close-up view of the active site. Closed form of loop 6 is from the 8RUC structure. Active site residues and RuBP substrate are labelled.






Update of
-
A map of the rubisco biochemical landscape.bioRxiv [Preprint]. 2024 Apr 11:2023.09.27.559826. doi: 10.1101/2023.09.27.559826. bioRxiv. 2024. Update in: Nature. 2025 Feb;638(8051):823-828. doi: 10.1038/s41586-024-08455-0. PMID: 38645011 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
