

The conformational landscape of human transthyretin revealed by cryo-EM
- PMID: 39843982
- PMCID: PMC12086048
- DOI: 10.1038/s41594-024-01472-7
The conformational landscape of human transthyretin revealed by cryo-EM
Abstract
Transthyretin (TTR) is a natively tetrameric thyroxine transporter in blood and cerebrospinal fluid whose misfolding and aggregation causes TTR amyloidosis. A rational drug design campaign identified the small molecule tafamidis (Vyndamax) as a stabilizer of the native TTR fold, and this aggregation inhibitor is regulatory agency approved for the treatment of TTR amyloidosis. Here we used cryo-EM to investigate the conformational landscape of this 55 kDa tetramer in the absence and presence of one or two ligands, revealing inherent asymmetries in the tetrameric architecture and previously unobserved conformational states. These findings provide critical mechanistic insights into negatively cooperative ligand binding and the structural pathways responsible for TTR amyloidogenesis, underscoring the capacity of cryo-EM to identify pharmacological targets suppressed by the confines of the crystal lattice, opening uncharted territory in structure-based drug design.
© 2025. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests: J.W.K. and E.T.P. discovered tafamidis and receive royalty payments for its sales. J.W.K. was a founder and shareholder in FoldRx, which first developed tafamidis as a therapeutic. J.W.K. is a paid consultant for and has received support for travel and accommodations from Pfizer, which sells tafamidis. The other authors declare no competing interests.
Figures




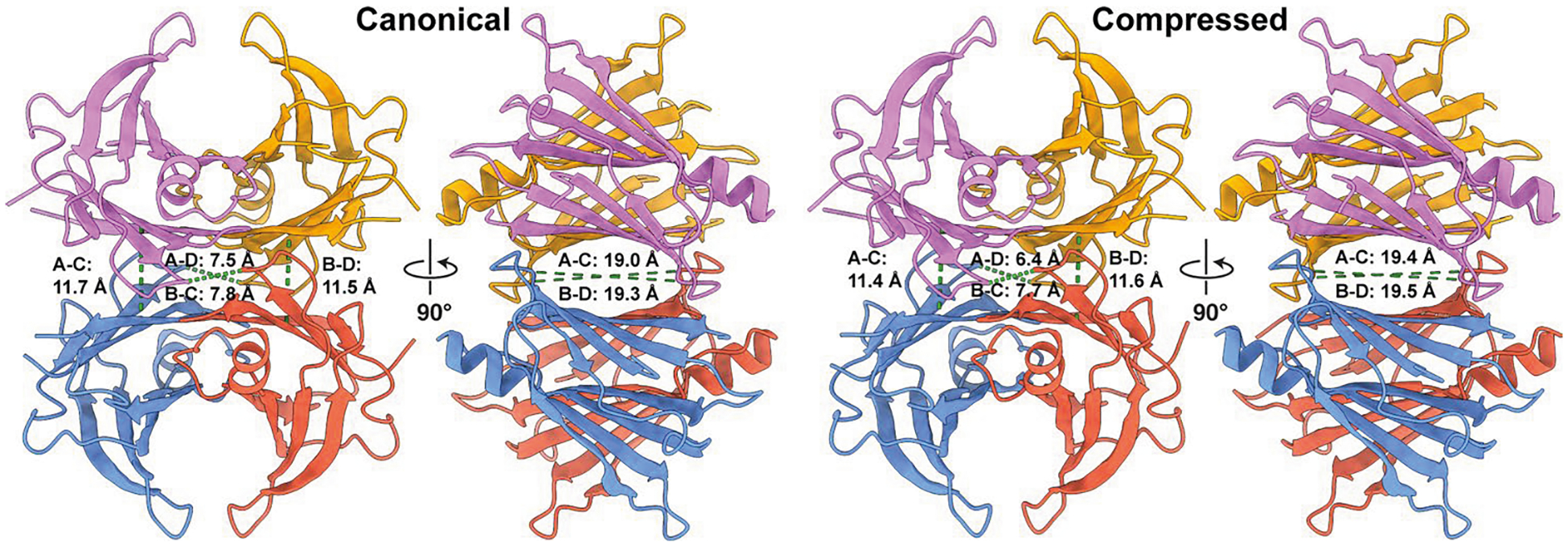


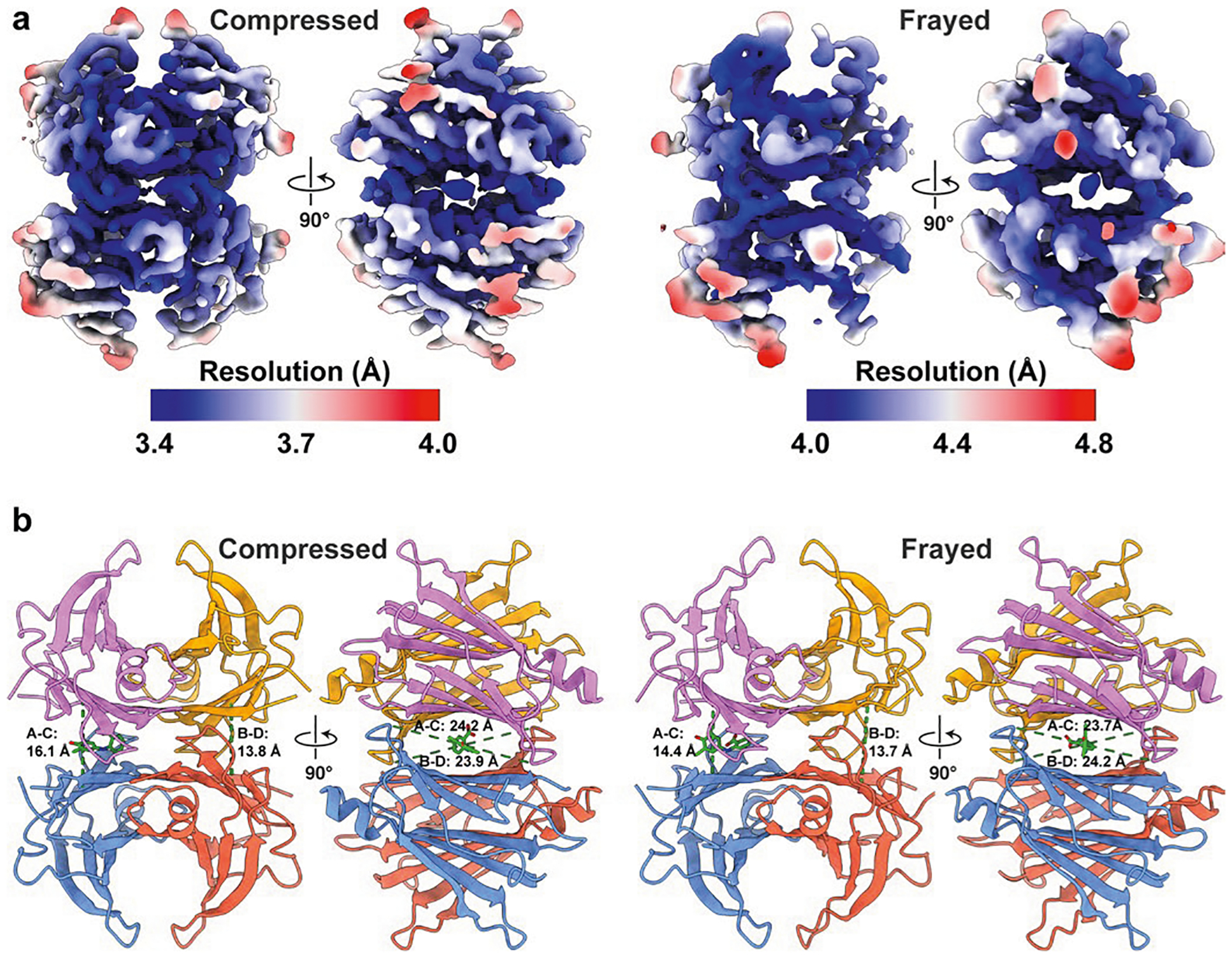






Update of
-
The conformational landscape of human transthyretin revealed by cryo-EM.bioRxiv [Preprint]. 2024 Jan 23:2024.01.23.576879. doi: 10.1101/2024.01.23.576879. bioRxiv. 2024. Update in: Nat Struct Mol Biol. 2025 May;32(5):876-883. doi: 10.1038/s41594-024-01472-7. PMID: 38328110 Free PMC article. Updated. Preprint.
Similar articles
-
The conformational landscape of human transthyretin revealed by cryo-EM.bioRxiv [Preprint]. 2024 Jan 23:2024.01.23.576879. doi: 10.1101/2024.01.23.576879. bioRxiv. 2024. Update in: Nat Struct Mol Biol. 2025 May;32(5):876-883. doi: 10.1038/s41594-024-01472-7. PMID: 38328110 Free PMC article. Updated. Preprint.
-
Repurposing Benzbromarone for Familial Amyloid Polyneuropathy: A New Transthyretin Tetramer Stabilizer.Int J Mol Sci. 2020 Sep 28;21(19):7166. doi: 10.3390/ijms21197166. Int J Mol Sci. 2020. PMID: 32998442 Free PMC article.
-
Tafamidis, a potent and selective transthyretin kinetic stabilizer that inhibits the amyloid cascade.Proc Natl Acad Sci U S A. 2012 Jun 12;109(24):9629-34. doi: 10.1073/pnas.1121005109. Epub 2012 May 29. Proc Natl Acad Sci U S A. 2012. PMID: 22645360 Free PMC article.
-
Nearly 200 X-ray crystal structures of transthyretin: what do they tell us about this protein and the design of drugs for TTR amyloidoses?Curr Med Chem. 2012;19(15):2324-42. doi: 10.2174/092986712800269335. Curr Med Chem. 2012. PMID: 22471981 Review.
-
A Comprehensive Review on Chemistry and Biology of Tafamidis in Transthyretin Amyloidosis.Mini Rev Med Chem. 2024;24(6):571-587. doi: 10.2174/0113895575241556231003055323. Mini Rev Med Chem. 2024. PMID: 37828667 Review.
Cited by
-
Differentiating the solution structures and stability of transthyretin tetramer complexed with tolcapone and tafamidis using SEC-SWAXS and NMR.J Appl Crystallogr. 2025 Jul 8;58(Pt 4):1373-1383. doi: 10.1107/S1600576725004716. eCollection 2025 Aug 1. J Appl Crystallogr. 2025. PMID: 40765976 Free PMC article.
-
Advances in uncovering the mechanisms of macromolecular conformational entropy.Nat Chem Biol. 2025 May;21(5):623-634. doi: 10.1038/s41589-025-01879-3. Epub 2025 Apr 24. Nat Chem Biol. 2025. PMID: 40275100 Free PMC article. Review.
-
"Proteinjury": a universal pathological mechanism mediated by cerebrospinal fluid in neurodegeneration and trauma.Front Cell Dev Biol. 2025 May 20;13:1593122. doi: 10.3389/fcell.2025.1593122. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40463840 Free PMC article. Review.
-
Sub-3 Å resolution protein structure determination by single-particle cryo-EM at 100 keV.Structure. 2025 Jul 26:S0969-2126(25)00255-2. doi: 10.1016/j.str.2025.07.007. Online ahead of print. Structure. 2025. PMID: 40744008
-
CryoSift - An accessible and automated CNN-driven tool for cryo-EM 2D class selection.bioRxiv [Preprint]. 2025 Aug 1:2025.07.28.667259. doi: 10.1101/2025.07.28.667259. bioRxiv. 2025. PMID: 40766614 Free PMC article. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
