

Altered Atlas of Exercise-Responsive MicroRNAs Revealing miR-29a-3p Attacks Armored and Cold Tumors and Boosts Anti-B7-H3 Therapy
- PMID: 39845707
- PMCID: PMC11751204
- DOI: 10.34133/research.0590
Altered Atlas of Exercise-Responsive MicroRNAs Revealing miR-29a-3p Attacks Armored and Cold Tumors and Boosts Anti-B7-H3 Therapy
Abstract
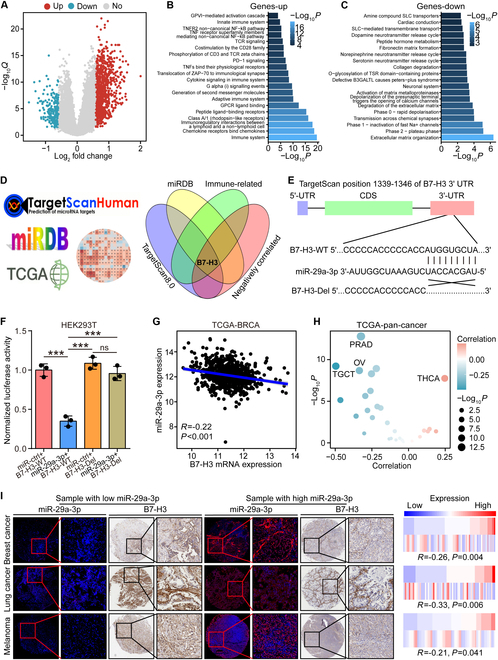
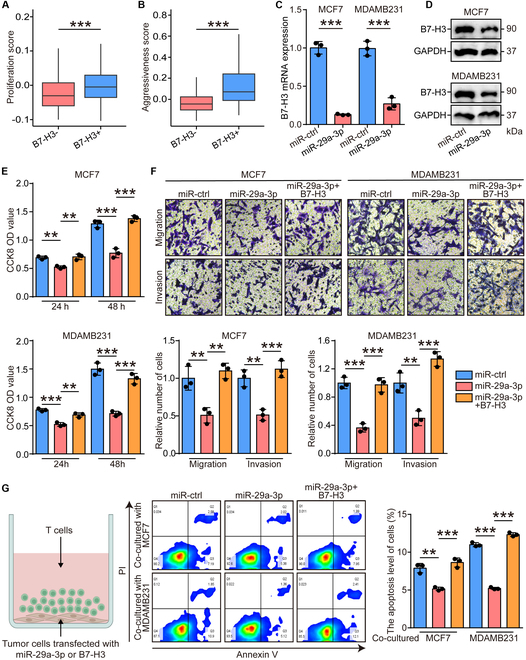
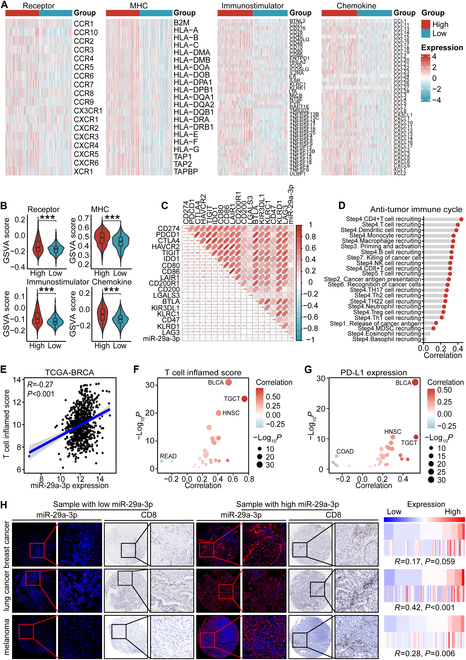
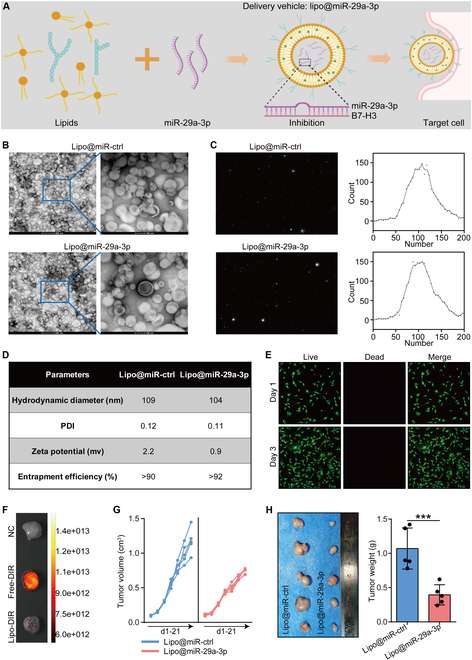
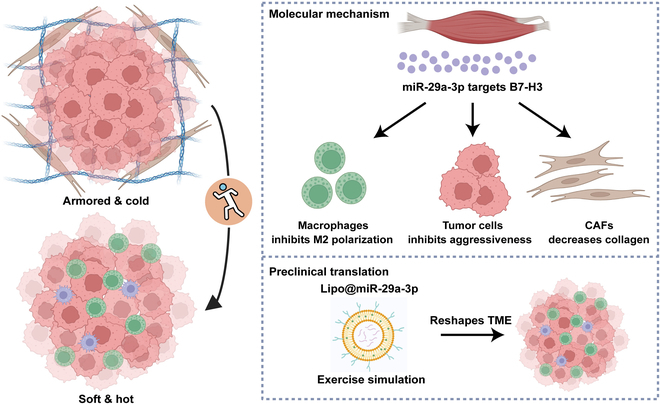
Increasing evidence has shown that physical exercise remarkably inhibits oncogenesis and progression of numerous cancers and exercise-responsive microRNAs (miRNAs) exert a marked role in exercise-mediated tumor suppression. In this research, expression and prognostic values of exercise-responsive miRNAs were examined in breast cancer (BRCA) and further pan-cancer types. In addition, multiple independent public and in-house cohorts, in vitro assays involving multiple, macrophages, fibroblasts, and tumor cells, and in vivo models were utilized to uncover the tumor-suppressive roles of miR-29a-3p in cancers. Here, we reported that miR-29a-3p was the exercise-responsive miRNA, which was lowly expressed in tumor tissues and associated with unfavorable prognosis in BRCA. Mechanistically, miR-29a-3p targeted macrophages, fibroblasts, and tumor cells to down-regulate B7 homolog 3 (B7-H3) expression. Single-cell RNA sequencing (scRNA-seq) and cytometry by time-of-flight (CyTOF) demonstrated that miR-29a-3p attacked the armored and cold tumors, thereby shaping an immuno-hot tumor microenvironment (TME). Translationally, liposomes were developed and loaded with miR-29a-3p (lipo@miR-29a-3p), and lipo@miR-29a-3p exhibited promising antitumor effects in a mouse model with great biocompatibility. In conclusion, we uncovered that miR-29a-3p is a critical exercise-responsive miRNA, which attacked armored and cold tumors by inhibiting B7-H3 expression. Thus, miR-29a-3p restoration could be an alternative strategy for antitumor therapy.
Copyright © 2025 Jie Mei et al.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.
Figures




Similar articles
-
MicroRNA miR-29a Inhibits Colon Cancer Progression by Downregulating B7-H3 Expression: Potential Molecular Targets for Colon Cancer Therapy.Mol Biotechnol. 2021 Sep;63(9):849-861. doi: 10.1007/s12033-021-00348-1. Epub 2021 Jun 7. Mol Biotechnol. 2021. PMID: 34100183
-
Voluntary exercise sensitizes cancer immunotherapy via the collagen inhibition-orchestrated inflammatory tumor immune microenvironment.Cell Rep. 2024 Sep 24;43(9):114697. doi: 10.1016/j.celrep.2024.114697. Epub 2024 Aug 31. Cell Rep. 2024. PMID: 39217611
-
Reduced miR-29a-3p expression is linked to the cell proliferation and cell migration in gastric cancer.World J Surg Oncol. 2015 Mar 12;13:101. doi: 10.1186/s12957-015-0513-x. World J Surg Oncol. 2015. PMID: 25889078 Free PMC article.
-
MiR-29a-3p suppresses cell proliferation in laryngocarcinoma by targeting prominin 1.FEBS Open Bio. 2017 Mar 18;7(5):645-651. doi: 10.1002/2211-5463.12199. eCollection 2017 May. FEBS Open Bio. 2017. PMID: 28469977 Free PMC article.
-
MiR-29a-3p: a potential biomarker and therapeutic target in colorectal cancer.Clin Transl Oncol. 2023 Mar;25(3):563-577. doi: 10.1007/s12094-022-02978-6. Epub 2022 Nov 10. Clin Transl Oncol. 2023. PMID: 36355327 Free PMC article. Review.
Cited by
-
Dual-faced CXCL5 holds the key to unlocking immunotherapy in obese pancreatic cancer.J Immunother Cancer. 2025 Aug 27;13(8):e012566. doi: 10.1136/jitc-2025-012566. J Immunother Cancer. 2025. PMID: 40866290 Free PMC article.
-
Adipose-derived small extracellular vesicle miR-146a-5p targets Fbx32 to regulate mitochondrial autophagy and delay aging in skeletal muscle.J Nanobiotechnology. 2025 Apr 10;23(1):287. doi: 10.1186/s12951-025-03367-1. J Nanobiotechnology. 2025. PMID: 40211295 Free PMC article.
-
MicroRNAs at the crossroads of exercise and ferroptosis: a regulatory bridge.Clin Exp Med. 2025 Jul 6;25(1):234. doi: 10.1007/s10238-025-01766-0. Clin Exp Med. 2025. PMID: 40618320 Free PMC article. Review.
References
-
- Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12–49. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
