

An amplification mechanism for weak ELF magnetic fields quantum-bio effects in cancer cells
- PMID: 39849096
- PMCID: PMC11757740
- DOI: 10.1038/s41598-025-87235-w
An amplification mechanism for weak ELF magnetic fields quantum-bio effects in cancer cells
Abstract
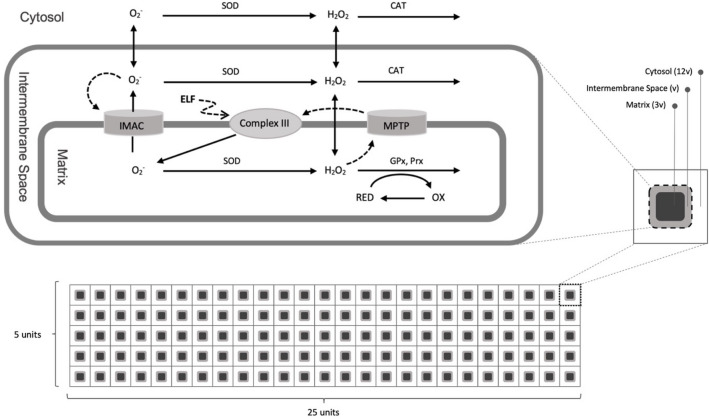
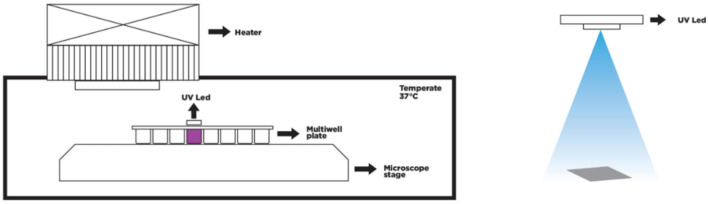
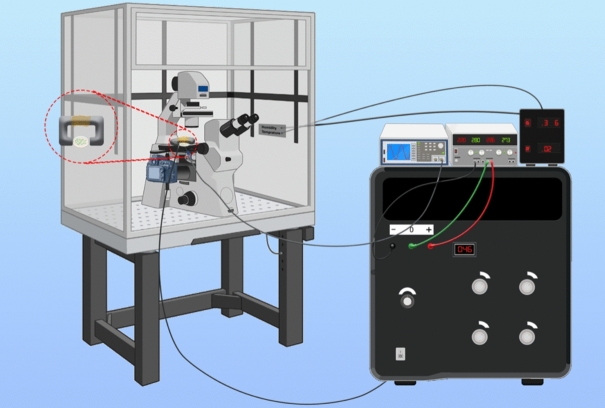
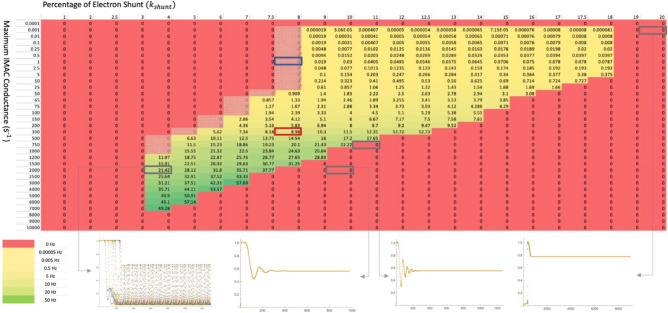
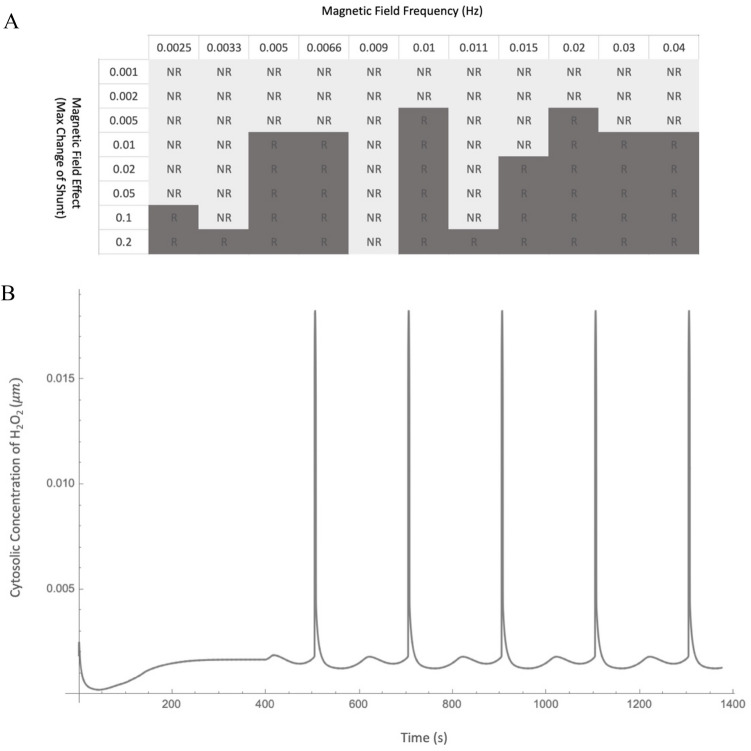
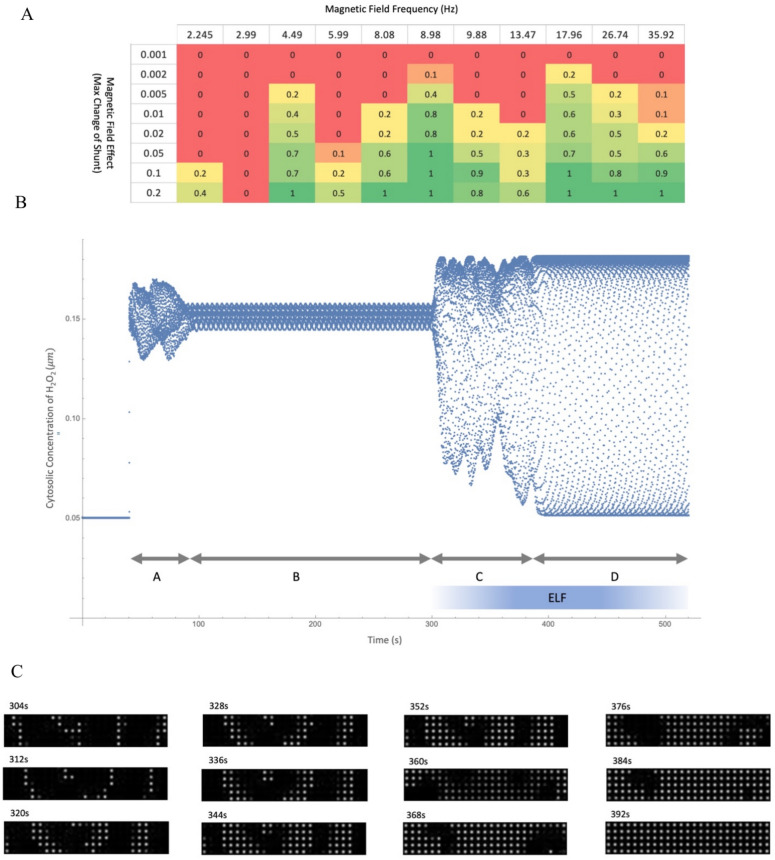
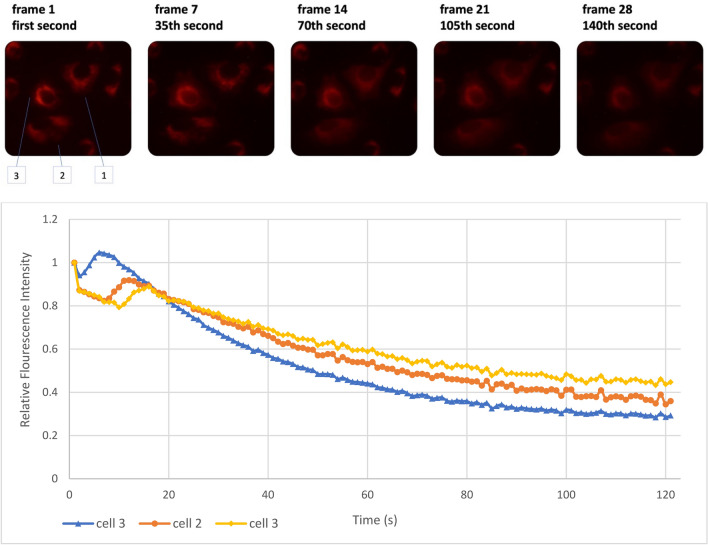
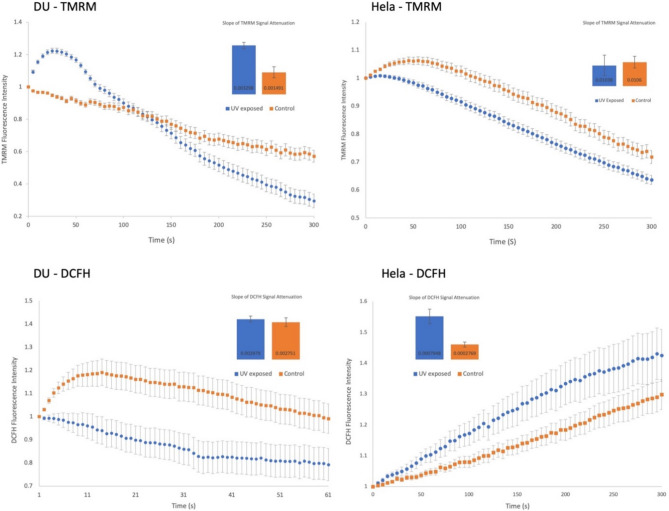
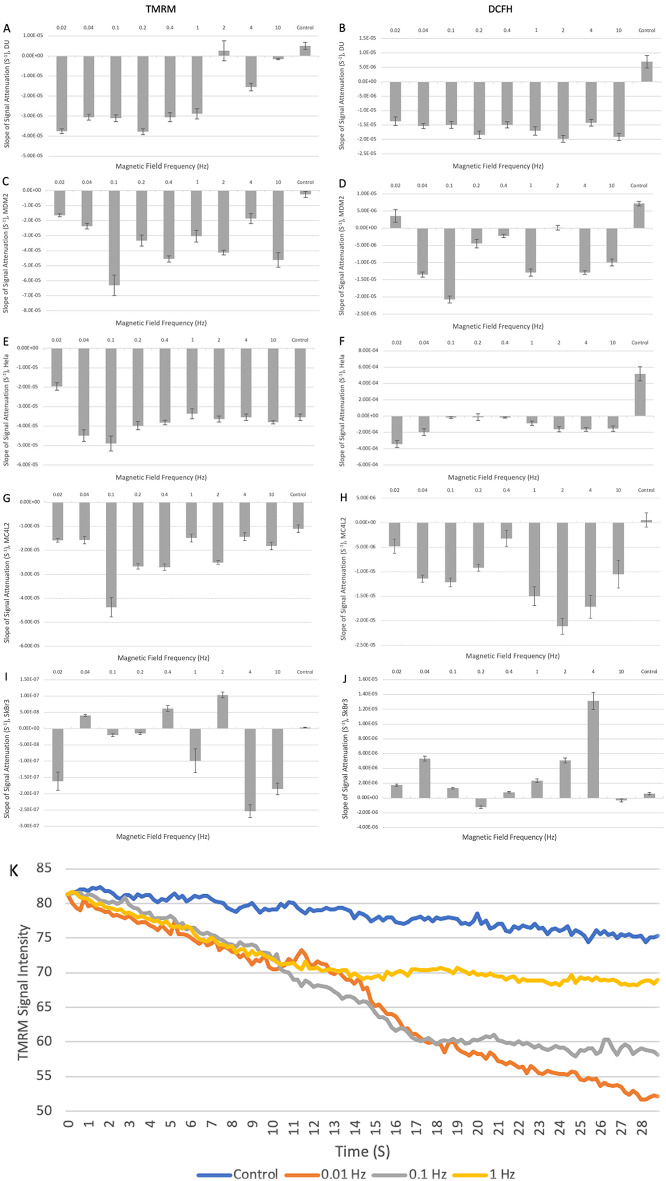
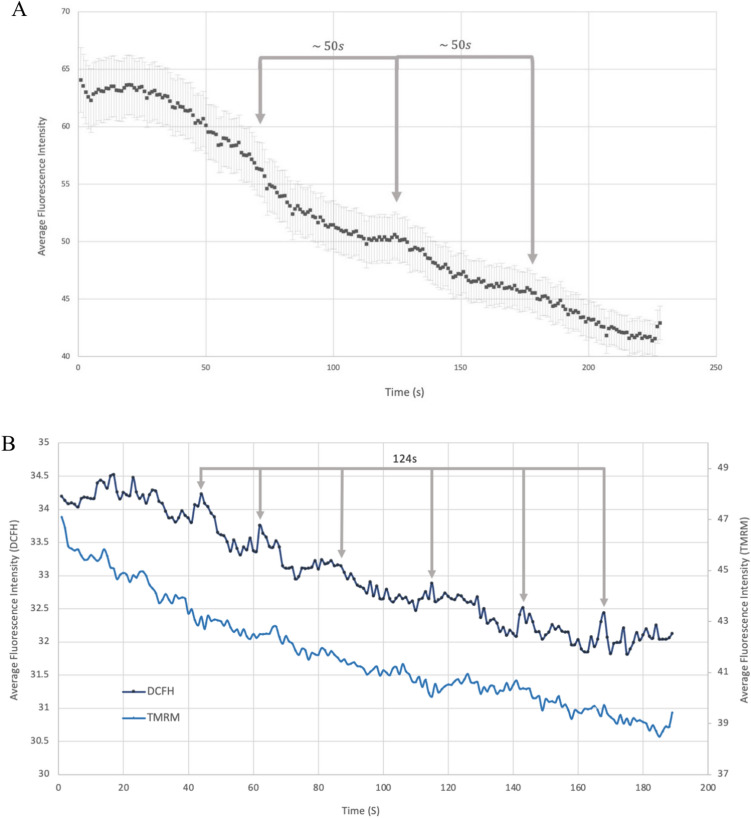
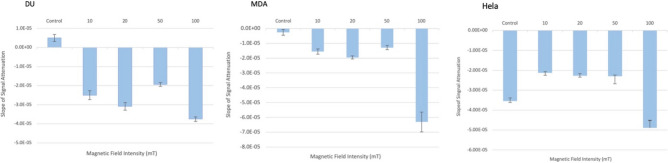
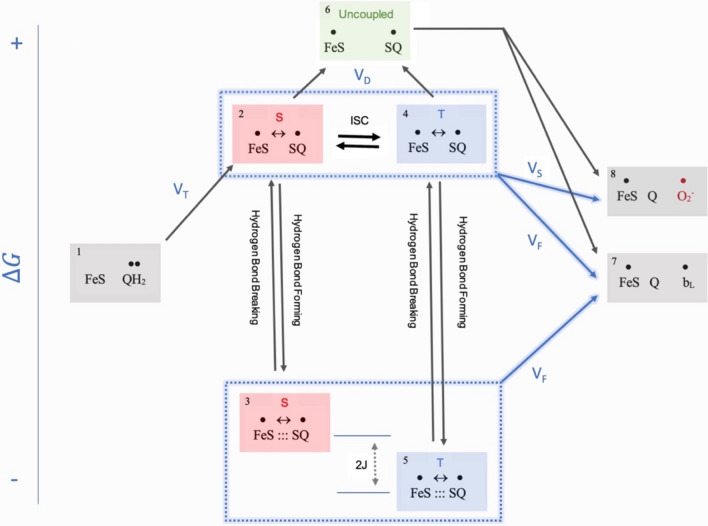
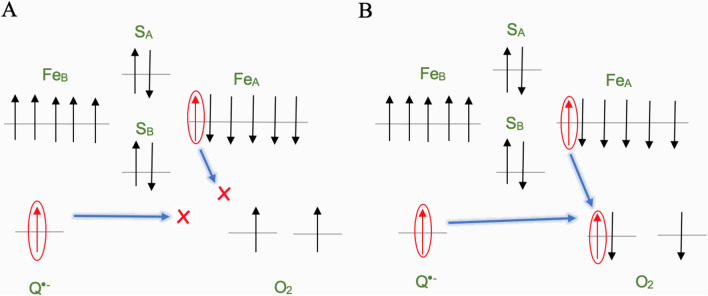
Observing quantum mechanical characteristics in biological processes is a surprising and important discovery. One example, which is gaining more experimental evidence and practical applications, is the effect of weak magnetic fields with extremely low frequencies on cells, especially cancerous ones. In this study, we use a mathematical model of ROS dynamics in cancer cells to show how ROS oscillatory patterns can act as a resonator to amplify the small effects of the magnetic fields on the radical pair dynamics in mitochondrial Complex III. We suggest such a resonator can act in two modes for distinct states in cancer cells: (1) cells at the edge of mitochondrial oscillation and (2) cells with local oscillatory patches. When exposed to magnetic fields, the first group exhibits high-amplitude oscillations, while the second group synchronizes to reach a whole-cell oscillation. Both types of amplification are frequency-dependent in the range of hertz and sub-hertz. We use UV radiation as a positive control to observe the two states of cells in DU and HELA cell lines. Application of magnetic fields shows frequency-dependent results on both the ROS and mitochondrial potential which agree with the model for both type of cells. We also observe the oscillatory behavior in the time-lapse fluorescence microscopy for 0.02 and 0.04 Hz magnetic fields. Finally, we investigate the dependence of the results on the field strength and propose a quantum spin-forbidden mechanism for the effect of magnetic fields on superoxide production in QO site of mitochondrial Complex III.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures


Similar articles
-
The effects of weak magnetic fields on radical pairs.Bioelectromagnetics. 2015 Jan;36(1):45-54. doi: 10.1002/bem.21883. Epub 2014 Nov 15. Bioelectromagnetics. 2015. PMID: 25399679
-
Induction of micronuclei and superoxide production in neuroblastoma and glioma cell lines exposed to weak 50 Hz magnetic fields.J R Soc Interface. 2016 Jan;13(114):20150995. doi: 10.1098/rsif.2015.0995. J R Soc Interface. 2016. PMID: 26791000 Free PMC article.
-
Effects of extremely low-frequency magnetic fields on human MDA-MB-231 breast cancer cells: proteomic characterization.Ecotoxicol Environ Saf. 2023 Mar 15;253:114650. doi: 10.1016/j.ecoenv.2023.114650. Epub 2023 Feb 16. Ecotoxicol Environ Saf. 2023. PMID: 36805133
-
Magnetocarcinogenesis: is there a mechanism for carcinogenic effects of weak magnetic fields?Proc Biol Sci. 2018 May 30;285(1879):20180590. doi: 10.1098/rspb.2018.0590. Proc Biol Sci. 2018. PMID: 29794049 Free PMC article. Review.
-
Molecular Biological Effects of Weak Low-Frequency Magnetic Fields: Frequency-Amplitude Efficiency Windows and Possible Mechanisms.Int J Mol Sci. 2023 Jul 1;24(13):10989. doi: 10.3390/ijms241310989. Int J Mol Sci. 2023. PMID: 37446167 Free PMC article. Review.
Cited by
-
Magneto-oncology: a radical pair primer.Front Oncol. 2025 Mar 7;15:1539718. doi: 10.3389/fonc.2025.1539718. eCollection 2025. Front Oncol. 2025. PMID: 40123899 Free PMC article. Review.
References
-
- Blackman, C. F. et al. Induction of calcium-ion efflux from brain tissue by radio-frequency radiation: Effects of modulation frequency and field strength. Radio Sci.14(6S), 93–98 (1979).
-
- Goodman, R., Bassett, C. A. & Henderson, A. S. Pulsing electromagnetic fields induce cellular transcription. Science220(4603), 1283–1285 (1983). - PubMed
-
- Reiter, R. J. Melatonin in the context of the reported bioeffects of environmental electromagnetic fields. Bioelectrochem. Bioenerg.47(1), 135–142 (1998).
-
- Adair, R. K. Constraints on biological effects of weak extremely-low-frequency electromagnetic fields. Phys. Rev. A43(2), 1039 (1991). - PubMed
-
- Binhi, V. N. & Rubin, A. B. Magnetobiology: the kT paradox and possible solutions. Electromagn. Biol. Med.26(1), 45–62 (2007). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
