

Macromolecular interactions and geometrical confinement determine the 3D diffusion of ribosome-sized particles in live Escherichia coli cells
- PMID: 39854229
- PMCID: PMC11789073
- DOI: 10.1073/pnas.2406340121
Macromolecular interactions and geometrical confinement determine the 3D diffusion of ribosome-sized particles in live Escherichia coli cells
Abstract
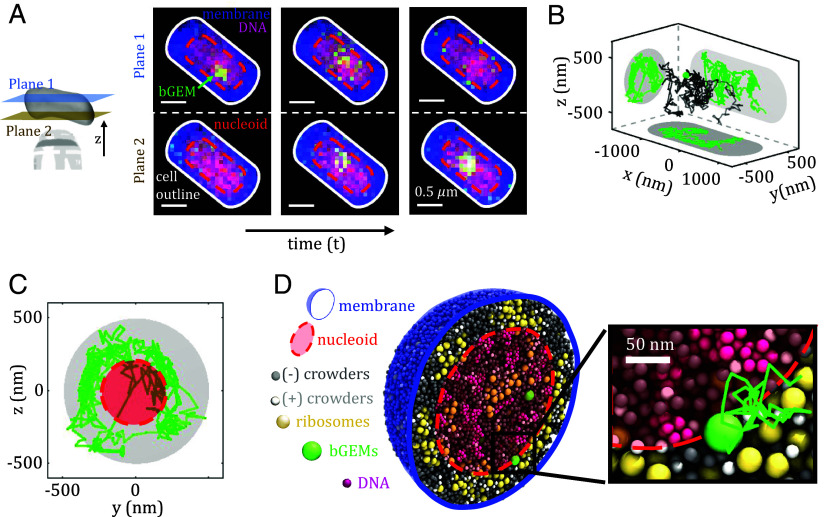
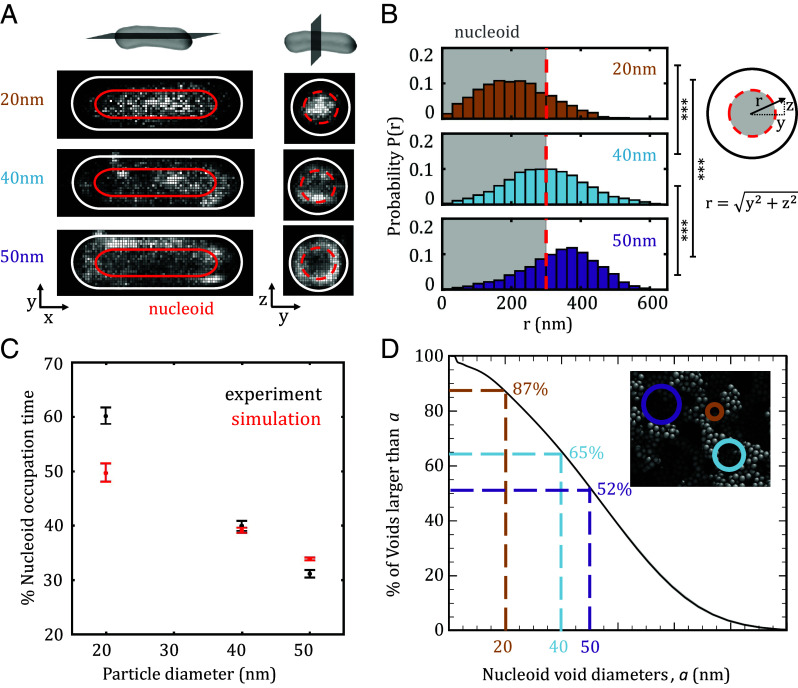
The crowded bacterial cytoplasm is composed of biomolecules that span several orders of magnitude in size and electrical charge. This complexity has been proposed as the source of the rich spatial organization and apparent anomalous diffusion of intracellular components, although this has not been tested directly. Here, we use biplane microscopy to track the 3D motion of self-assembled bacterial genetically encoded multimeric nanoparticles (bGEMs) with tunable size (20 to 50 nm) and charge (-3,240 to +2,700 e) in live Escherichia coli cells. To probe intermolecular details at spatial and temporal resolutions beyond experimental limits, we also developed a colloidal whole-cell model that explicitly represents the size and charge of cytoplasmic macromolecules and the porous structure of the bacterial nucleoid. Combining these techniques, we show that bGEMs spatially segregate by size, with small 20-nm particles enriched inside the nucleoid, and larger and/or positively charged particles excluded from this region. Localization is driven by entropic and electrostatic forces arising from cytoplasmic polydispersity, nucleoid structure, geometrical confinement, and interactions with other biomolecules including ribosomes and DNA. We observe that at the timescales of traditional single molecule tracking experiments, motion appears subdiffusive for all particle sizes and charges. However, using computer simulations with higher temporal resolution, we find that the apparent anomalous exponents are governed by the region of the cell in which bGEMs are located. Molecular motion does not display anomalous diffusion on short time scales and the apparent subdiffusion arises from geometrical confinement within the nucleoid and by the cell boundary.
Keywords: anomalous diffusion; bacterial cytoplasm; biophysics; microbiology.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures


Update of
-
Macromolecular interactions and geometrical confinement determine the 3D diffusion of ribosome-sized particles in live Escherichia coli cells.bioRxiv [Preprint]. 2024 Mar 28:2024.03.27.587083. doi: 10.1101/2024.03.27.587083. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2025 Jan 28;122(4):e2406340121. doi: 10.1073/pnas.2406340121. PMID: 38585850 Free PMC article. Updated. Preprint.
Similar articles
-
Macromolecular interactions and geometrical confinement determine the 3D diffusion of ribosome-sized particles in live Escherichia coli cells.bioRxiv [Preprint]. 2024 Mar 28:2024.03.27.587083. doi: 10.1101/2024.03.27.587083. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2025 Jan 28;122(4):e2406340121. doi: 10.1073/pnas.2406340121. PMID: 38585850 Free PMC article. Updated. Preprint.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Psychological interventions for adults who have sexually offended or are at risk of offending.Cochrane Database Syst Rev. 2012 Dec 12;12(12):CD007507. doi: 10.1002/14651858.CD007507.pub2. Cochrane Database Syst Rev. 2012. PMID: 23235646 Free PMC article.
-
Systemic Inflammatory Response Syndrome.2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31613449 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
Cited by
-
Single Particle Tracking of Genetically Encoded Nanoparticles: Optimizing Expression for Cytoplasmic Diffusion Studies.bioRxiv [Preprint]. 2024 Nov 18:2024.11.17.623896. doi: 10.1101/2024.11.17.623896. bioRxiv. 2024. Update in: Biophys J. 2025 Jul 1;124(13):2222-2235. doi: 10.1016/j.bpj.2025.05.025. PMID: 39605363 Free PMC article. Updated. Preprint.
-
Giant KASH proteins and ribosomes synergistically establish cytoplasmic biophysical properties in vivo.bioRxiv [Preprint]. 2025 Jan 12:2025.01.10.632479. doi: 10.1101/2025.01.10.632479. bioRxiv. 2025. PMID: 39829784 Free PMC article. Preprint.
References
-
- Mignot T., Shaevitz J. W., Active and passive mechanisms of intracellular transport and localization in bacteria. Curr. Opin. Microbiol. 11, 580–585 (2008). - PubMed
MeSH terms
Substances
Grants and funding
- RGP0016/2022-102/Human Frontier Science Program (HFSP)
- R37 CA240765/CA/NCI NIH HHS/United States
- TR01 NS127186/HHS | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- R01 NS127186/NS/NINDS NIH HHS/United States
- GRF 1656518/NSF | National Science Foundation Graduate Research Fellowship Program (GRFP)
LinkOut - more resources
Full Text Sources
