

This is a preprint.
Reconstructing NOD-like receptor alleles with high internal conservation in Podospora anserina using long-read sequencing
- PMID: 39868110
- PMCID: PMC11761791
- DOI: 10.1101/2025.01.13.632504
Reconstructing NOD-like receptor alleles with high internal conservation in Podospora anserina using long-read sequencing
Update in
-
Reconstructing NOD-like receptor alleles with high internal conservation in Podospora anserina using long-read sequencing.Microb Genom. 2025 Jul;11(7):001442. doi: 10.1099/mgen.0.001442. Microb Genom. 2025. PMID: 40601474 Free PMC article.
Abstract
NOD-like receptors (NLRs) are intracellular immune receptors that detect pathogen-associated cues and trigger defense mechanisms, including regulated cell death. In filamentous fungi, some NLRs mediate heterokaryon incompatibility, a self/non-self recognition process that prevents the vegetative fusion of genetically distinct individuals, reducing the risk of parasitism. The het-d and het-e NLRs in Podospora anserina are highly polymorphic incompatibility genes (het genes) whose products recognize different alleles of the het-c gene via a sensor domain composed of WD40 repeats. These repeats display unusually high sequence identity maintained by concerted evolution. However, some sites within individual repeats are hypervariable and under diversifying selection. Despite extensive genetic studies, inconsistencies in the reported WD40 domain sequence have hindered functional and evolutionary analyses. Here we demonstrate that the WD40 domain can be accurately reconstructed from long-read sequencing (Oxford Nanopore and PacBio) data, but not from Illumina-based assemblies. Functional alleles are usually formed by 11 highly conserved repeats, with different repeat combinations underlying the same phenotypic het-d and het-e incompatibility reactions. Protein structure models suggest that their WD40 domain folds into two 7-blade β-propellers composed of the highly conserved repeats, as well as three cryptic divergent repeats at the C-terminus. We additionally show that one particular het-e allele does not have an incompatibility reaction with common het-c alleles, despite being 11-repeats long. Our findings provide a robust foundation for future research into the molecular mechanisms and evolutionary dynamics of het NLRs, while also highlighting both the fragility and the flexibility of β-propellers as immune sensor domains.
Keywords: WD40 domain; allorecognition; fungi; heterokaryon incompatibility.
Figures
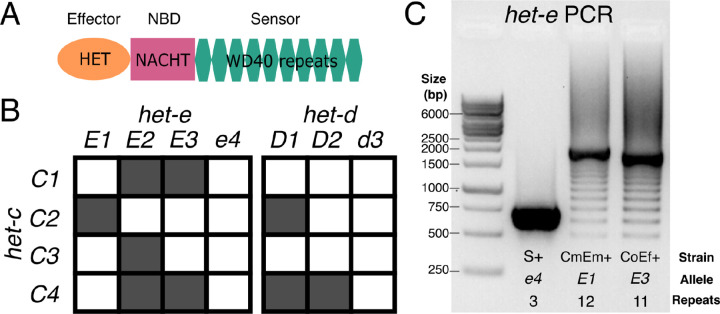
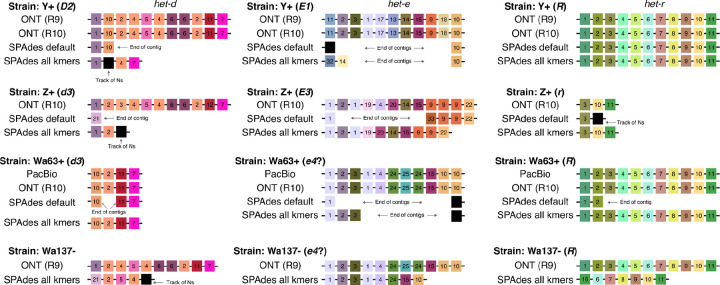


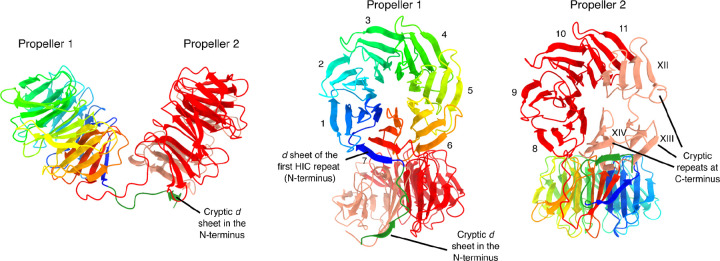
References
-
- Aanen DK, Debets AJM, De Visser JAGM, Hoekstra RF. 2008. The social evolution of somatic fusion. BioEssays 30: 1193–1203. - PubMed
-
- Allen RL, Bittner-Eddy PD, Grenville-Briggs LJ, Meitz JC, Rehmany AP, Rose LE, Beynon JL. 2004. Host-Parasite Coevolutionary Conflict Between Arabidopsis and Downy Mildew. Science 306: 1957–1960. - PubMed
Publication types
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources