

Fatty acid metabolism and the oxidative stress response support bacterial predation
- PMID: 39869799
- PMCID: PMC11804543
- DOI: 10.1073/pnas.2420875122
Fatty acid metabolism and the oxidative stress response support bacterial predation
Abstract
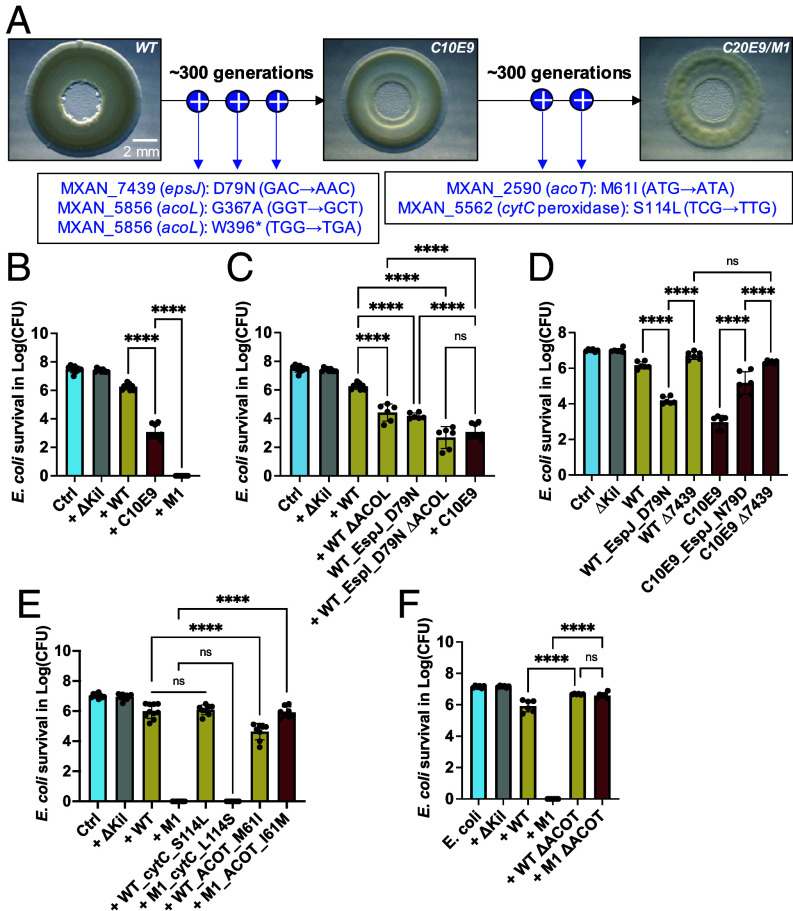
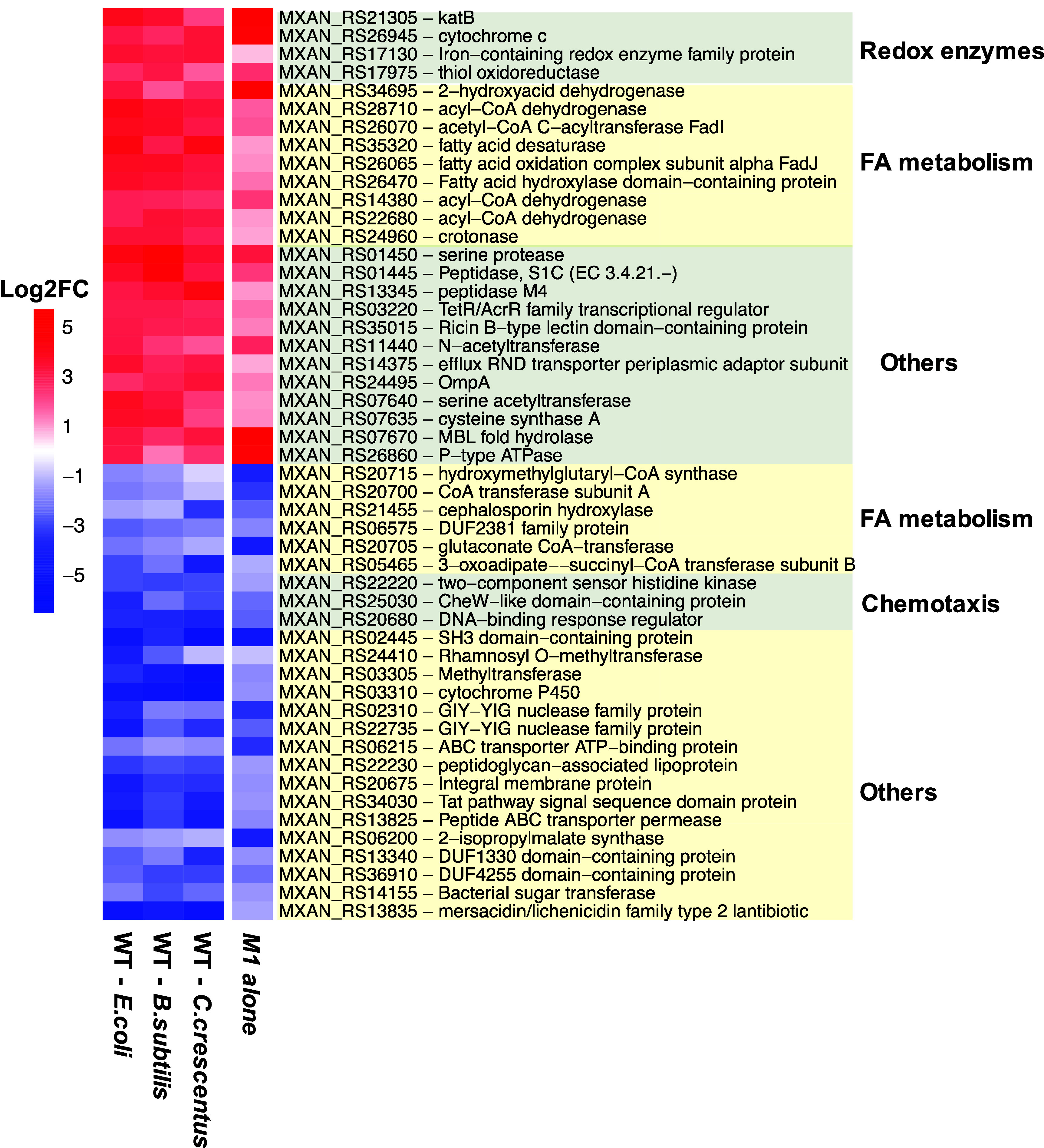
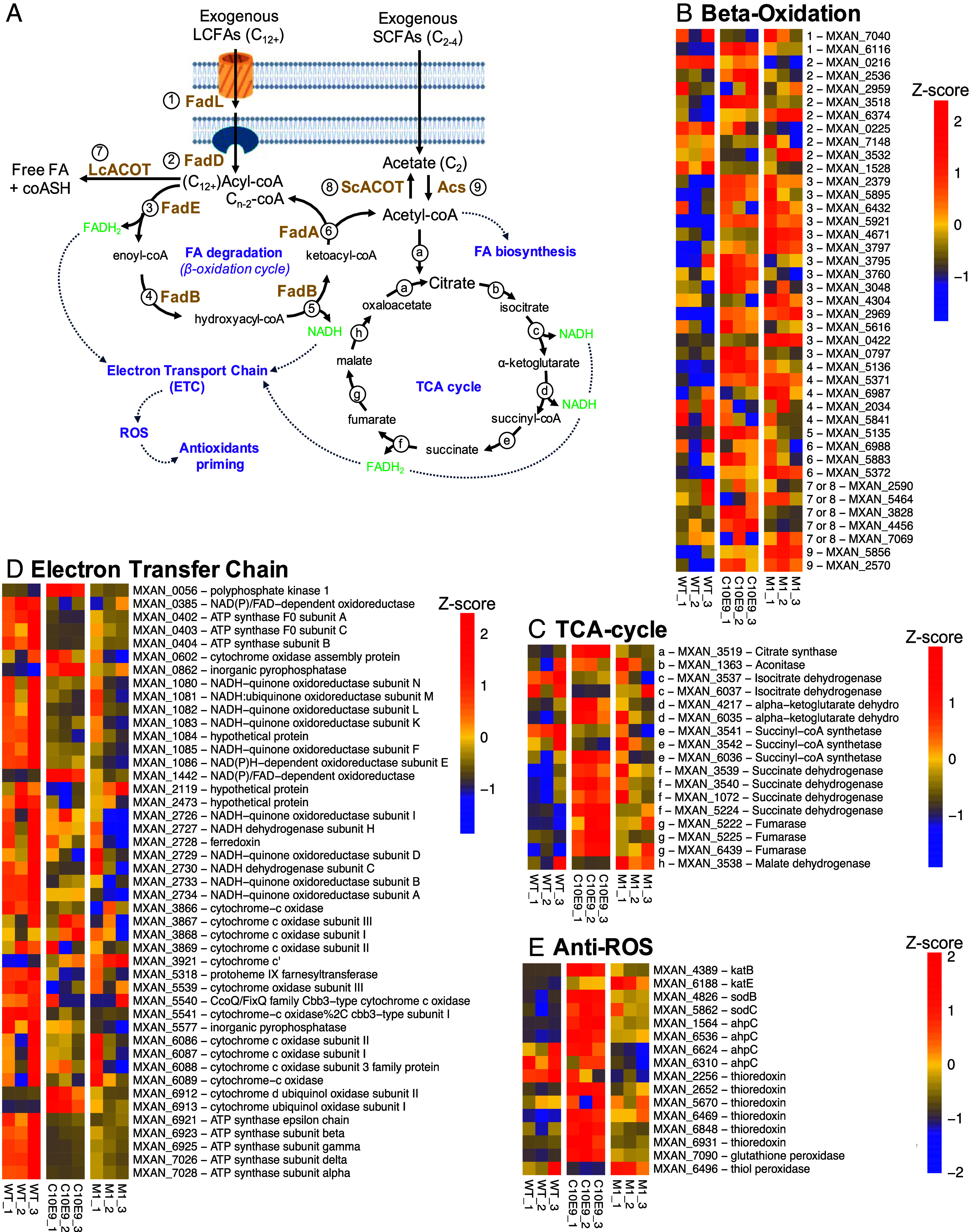
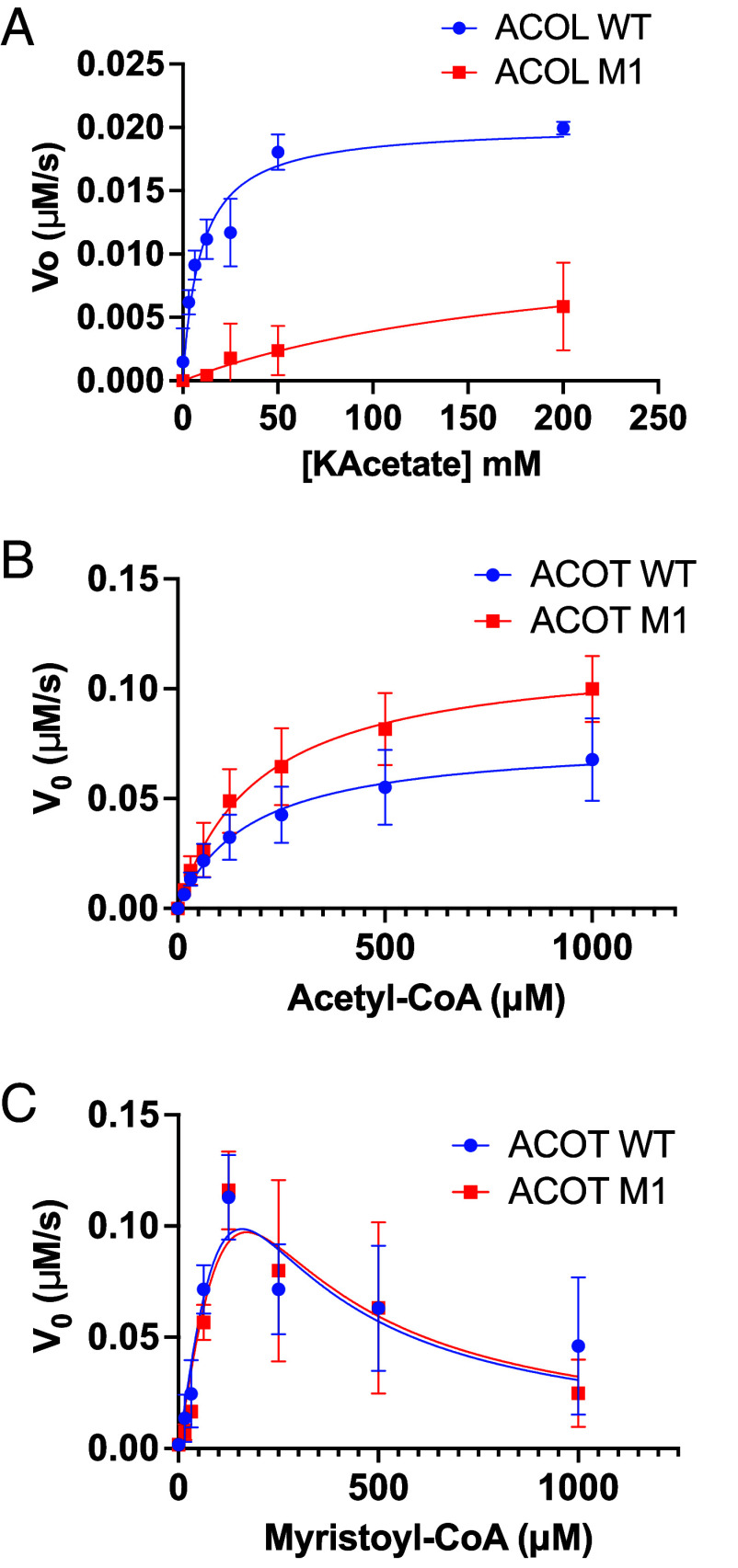
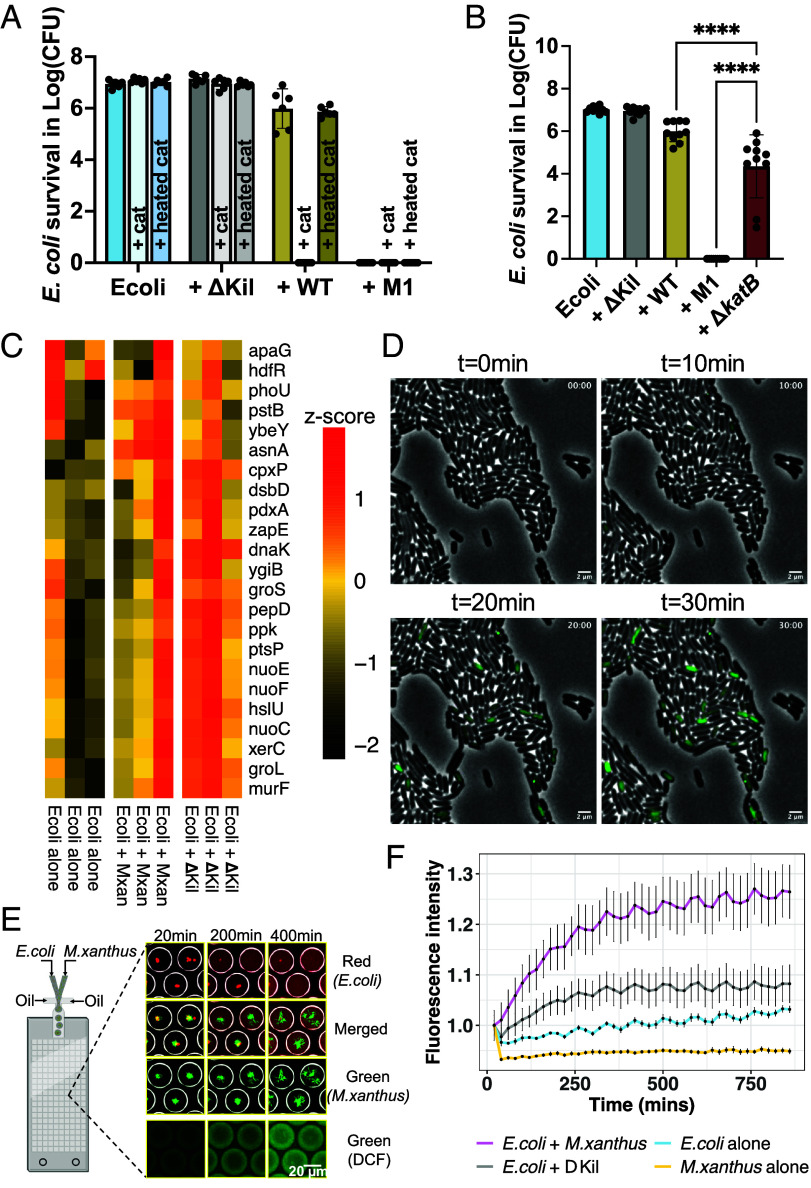
Despite growing awareness of their importance in soil ecology, the genetic and physiological traits of bacterial predators are still relatively poorly understood. In the course of a Myxococcus xanthus predator evolution experiment, we identified a class of genotypes leading to enhanced predation against diverse species. RNA-seq analysis demonstrated that this phenotype is linked to the constitutive activation of a predation-specific program. Functional analysis of the mutations accumulated across the evolutionary time in a two-component system and Acyl-CoA-manipulating enzymes revealed the critical roles of fatty acid metabolism and antioxidant gene induction. The former likely adapts the predator to metabolites derived from the prey while the latter protects predatory cells from reactive oxygen species generated by prey cells under stress and released upon lysis during predation. These findings reveal interesting parallels between bacterial predator-prey dynamics and pathogen-host cell interactions.
Keywords: Myxococcus xanthus; bacterial predation; experimental evolution; fatty acid metabolism; oxidative stress.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures


Similar articles
-
DdiA, an XRE family transcriptional regulator, is a co-regulator of the DNA damage response in Myxococcus xanthus.J Bacteriol. 2025 Jul 24;207(7):e0018425. doi: 10.1128/jb.00184-25. Epub 2025 Jul 3. J Bacteriol. 2025. PMID: 40608330 Free PMC article.
-
Experimental assessment of interactions between marine bacteria and model protists: from predator-prey relationships to bacterial-mediated lysis.Appl Environ Microbiol. 2025 Jun 18;91(6):e0092925. doi: 10.1128/aem.00929-25. Epub 2025 May 30. Appl Environ Microbiol. 2025. PMID: 40444979 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
A global assessment of large terrestrial carnivore kill rates.Biol Rev Camb Philos Soc. 2025 Feb;100(1):327-350. doi: 10.1111/brv.13143. Epub 2024 Sep 11. Biol Rev Camb Philos Soc. 2025. PMID: 39262094 Free PMC article.
-
Milestones in the development of Myxococcus xanthus as a model multicellular bacterium.J Bacteriol. 2025 Jul 24;207(7):e0007125. doi: 10.1128/jb.00071-25. Epub 2025 Jun 17. J Bacteriol. 2025. PMID: 40525847 Free PMC article. Review.
Cited by
-
Tad pili with adaptable tips mediate contact-dependent killing during bacterial predation.Nat Commun. 2025 May 13;16(1):4425. doi: 10.1038/s41467-025-58967-0. Nat Commun. 2025. PMID: 40360469 Free PMC article.
References
-
- Meena R. S., et al. , Impact of agrochemicals on soil microbiota and management: A review. Land 9, 34 (2020).
-
- Seabrook L., Mcalpine C. A., Bowen M. E., Restore, repair or reinvent: Options for sustainable landscapes in a changing climate. Landsc. Urban Plan. 100, 407–410 (2011).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
