

RECQL4 requires PARP1 for recruitment to DNA damage, and PARG dePARylation facilitates its associated role in end joining
- PMID: 39870799
- PMCID: PMC11799438
- DOI: 10.1038/s12276-024-01383-z
RECQL4 requires PARP1 for recruitment to DNA damage, and PARG dePARylation facilitates its associated role in end joining
Abstract
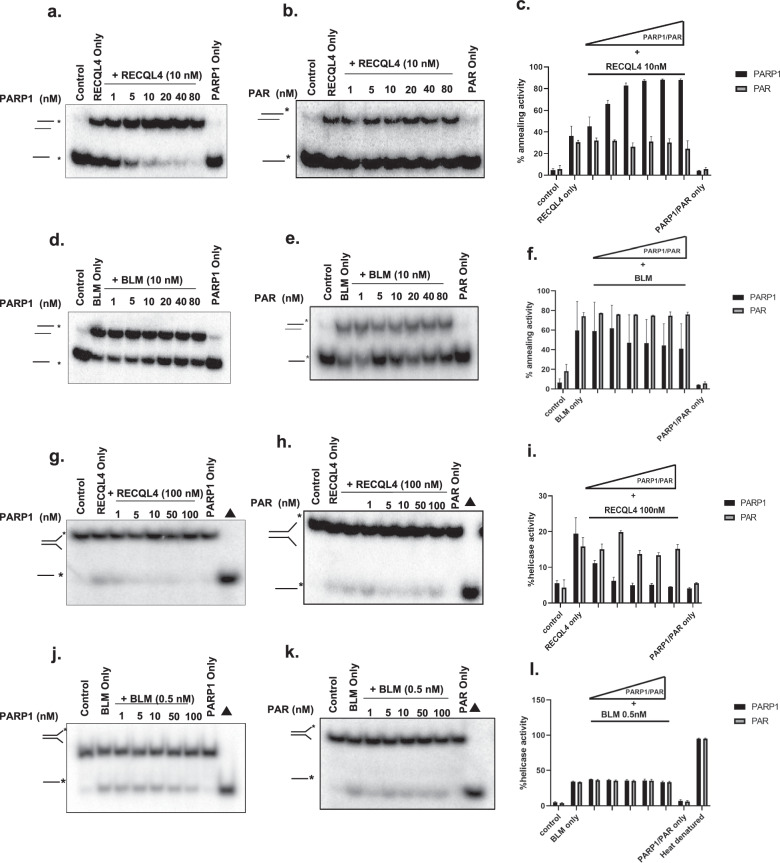
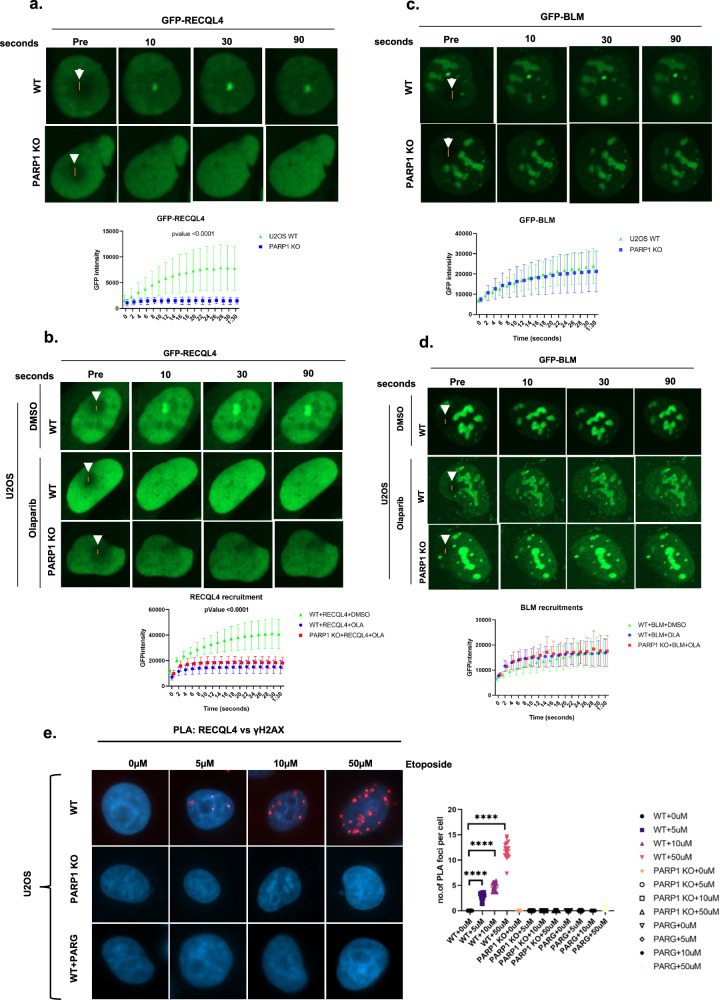
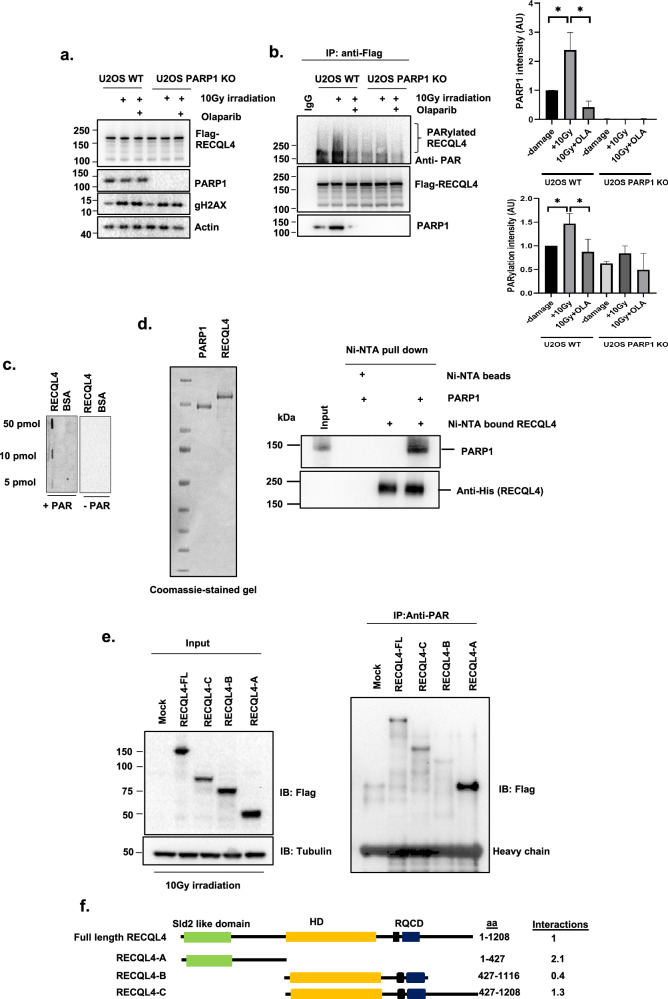
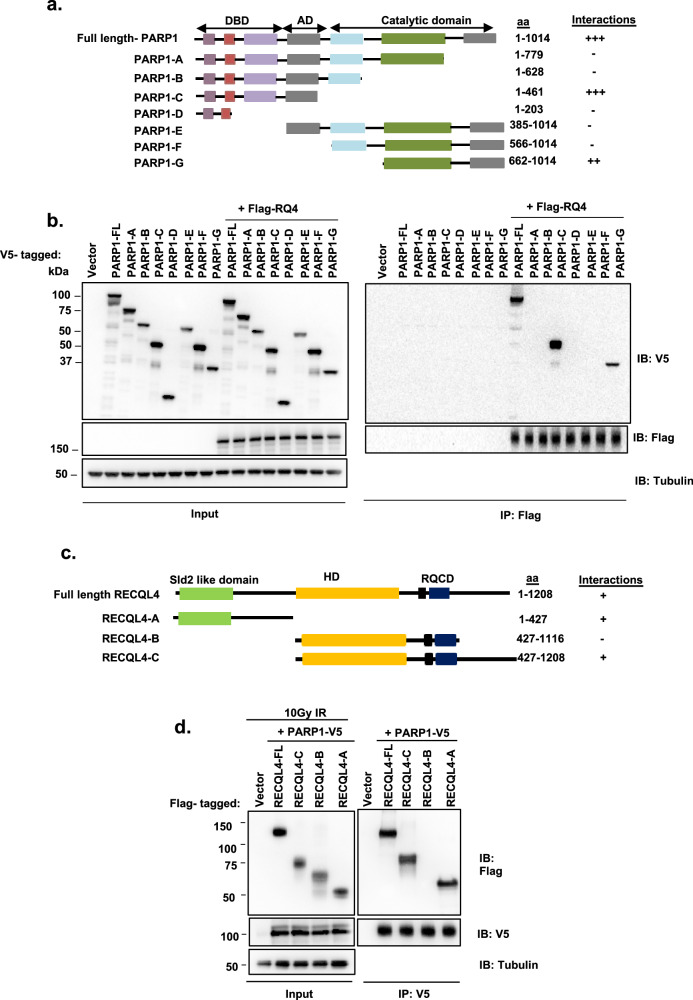
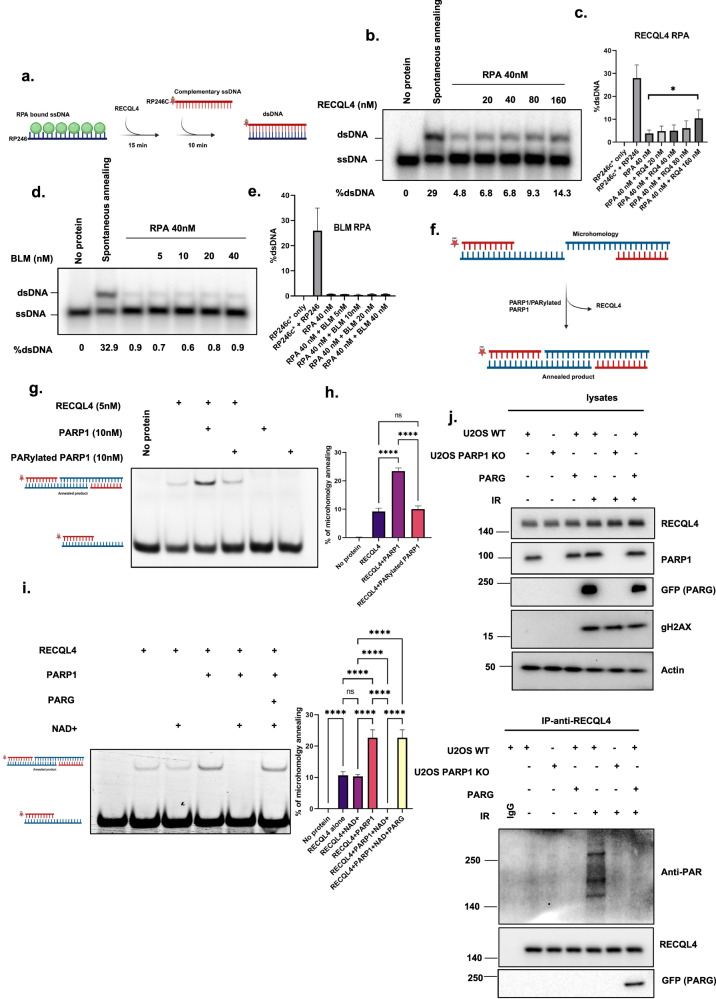
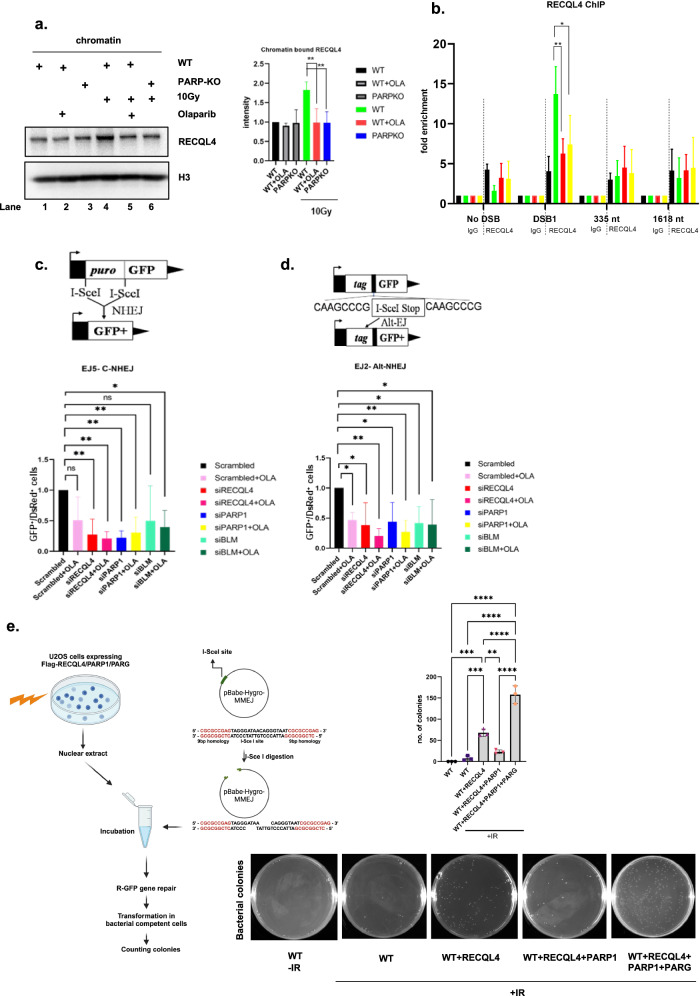
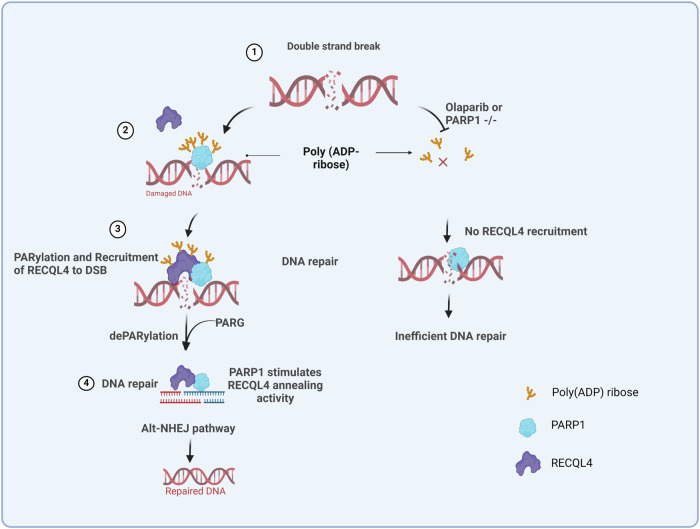
RecQ helicases, highly conserved proteins with pivotal roles in DNA replication, DNA repair and homologous recombination, are crucial for maintaining genomic integrity. Mutations in RECQL4 have been associated with various human diseases, including Rothmund-Thomson syndrome. RECQL4 is involved in regulating major DNA repair pathways, such as homologous recombination and nonhomologous end joining (NHEJ). RECQL4 has more prominent single-strand DNA annealing activity than helicase activity. Its ability to promote DNA damage repair and the precise role of its DNA annealing activity in DNA repair are unclear. Here we demonstrate that PARP1 interacts with RECQL4, increasing its single-stranded DNA strand annealing activity. PARP1 specifically promoted RECQL4 PARylation at both its N- and C-terminal regions, promoting RECQL4 recruitment to DNA double-strand breaks (DSBs). Inhibition or depletion of PARP1 significantly diminished RECQL4 recruitment and occupancy at specific DSB sites on chromosomes. After DNA damage, PARG dePARylated RECQL4 and stimulated its end-joining activity. RECQL4 actively displaced replication protein A from single-stranded DNA, promoting microhomology annealing in vitro. Furthermore, depletion of PARP1 or RECQL4 substantially impacted classical-NHEJ- and alternative-NHEJ-mediated DSB repair. Consequently, the combined activities of PARP1, PARG and RECQL4 modulate DNA repair.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
-
- Jeggo, P. A. & Lobrich, M. How cancer cells hijack DNA double-strand break repair pathways to gain genomic instability. Biochem. J.471, 1–11 (2015). - PubMed
-
- Vermeij, W. P., Hoeijmakers, J. H. & Pothof, J. Genome integrity in aging: human syndromes, mouse models, and therapeutic options. Annu. Rev. Pharmacol. Toxicol. 56, 427–445 (2016). - PubMed
-
- Srivastava, M. & Raghavan, S. C. DNA double-strand break repair inhibitors as cancer therapeutics. Chem. Biol.22, 17–29 (2015). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
