

DNA spontaneously wrapping around a histone core prefers negative supercoiling: A Brownian dynamics study
- PMID: 39874389
- PMCID: PMC11793753
- DOI: 10.1371/journal.pcbi.1012362
DNA spontaneously wrapping around a histone core prefers negative supercoiling: A Brownian dynamics study
Abstract
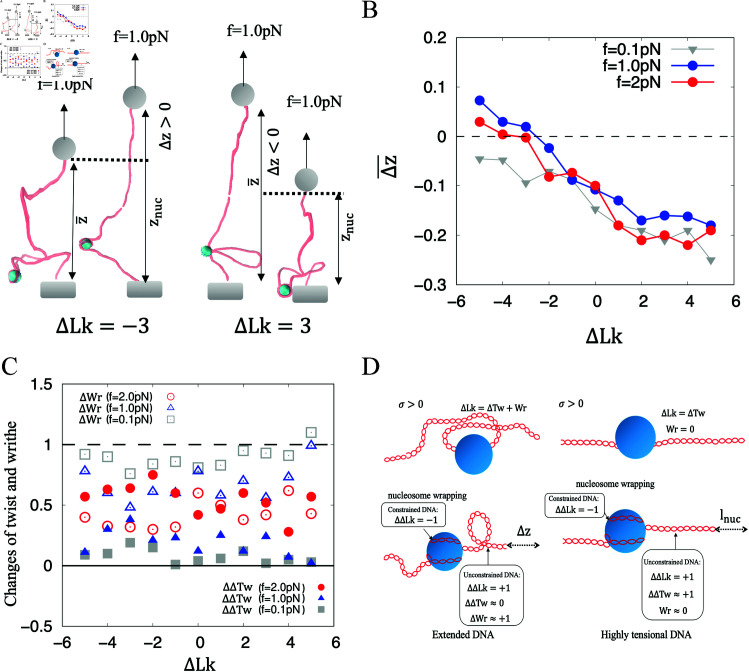
In eukaryotes, DNA achieves a highly compact structure primarily due to its winding around the histone cores. The nature wrapping of DNA around histone core form a 1.7 left-handed superhelical turns, contributing to negative supercoiling in chromatin. During transcription, negative supercoils generated behind the polymerase during transcription may play a role in triggering nucleosome reassembly. To elucidate how supercoils influence the dynamics of wrapping of DNA around the histone cores, we developed a novel model to simulate the intricate interplay between DNA and histone. Our simulations reveal that both positively and negatively supercoiled DNAs are capable of wrapping around histone cores to adopt the nucleosome conformation. Notably, our findings confirm a strong preference for negative supercoiled DNA during nucleosome wrapping, and reveal that the both of the negative writhe and twist are beneficial to the formation of the DNA wrapping around histone. Additionally, the simulations of the multiple nucleosomes on the same DNA template indicate that the nucleosome tends to assemble in proximity to the original nucleosome. This advancement in understanding the spontaneous formation of nucleosomes may offer insights into the complex dynamics of chromatin assembly and the fundamental mechanisms governing the structure and function of chromatin.
Copyright: © 2025 Long et al. This is an open access article distributed under the terms of the CreativeCommonsAttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
The supercoiling state of DNA determines the handedness of both H3 and CENP-A nucleosomes.Nanoscale. 2017 Feb 2;9(5):1862-1870. doi: 10.1039/c6nr06245h. Nanoscale. 2017. PMID: 28094382 Free PMC article.
-
Effect of positive supercoiling on DNA compaction by nucleosome cores.J Mol Biol. 1993 Nov 20;234(2):297-301. doi: 10.1006/jmbi.1993.1585. J Mol Biol. 1993. PMID: 8230214
-
Negative constrained DNA supercoiling in archaeal nucleosomes.Mol Microbiol. 2000 Jan;35(2):341-9. doi: 10.1046/j.1365-2958.2000.01689.x. Mol Microbiol. 2000. PMID: 10652094
-
Breaths, Twists, and Turns of Atomistic Nucleosomes.J Mol Biol. 2021 Mar 19;433(6):166744. doi: 10.1016/j.jmb.2020.166744. Epub 2020 Dec 10. J Mol Biol. 2021. PMID: 33309853 Review.
-
Chromatosome Structure and Dynamics from Molecular Simulations.Annu Rev Phys Chem. 2020 Apr 20;71:101-119. doi: 10.1146/annurev-physchem-071119-040043. Epub 2020 Feb 4. Annu Rev Phys Chem. 2020. PMID: 32017651 Review.
References
-
- Bates AD, Maxwell A, Maxwell T. DNA topology. Oxford: Oxford University Press; 2005.
-
- Kornberg RD. Structure of chromatin. Annu Rev Biochem. 2003:46(1);931. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
