

NFAT5 exacerbates β-cell ferroptosis by suppressing the transcription of PRDX2 in obese type 2 diabetes mellitus
- PMID: 39875646
- PMCID: PMC11775373
- DOI: 10.1007/s00018-024-05563-y
NFAT5 exacerbates β-cell ferroptosis by suppressing the transcription of PRDX2 in obese type 2 diabetes mellitus
Abstract
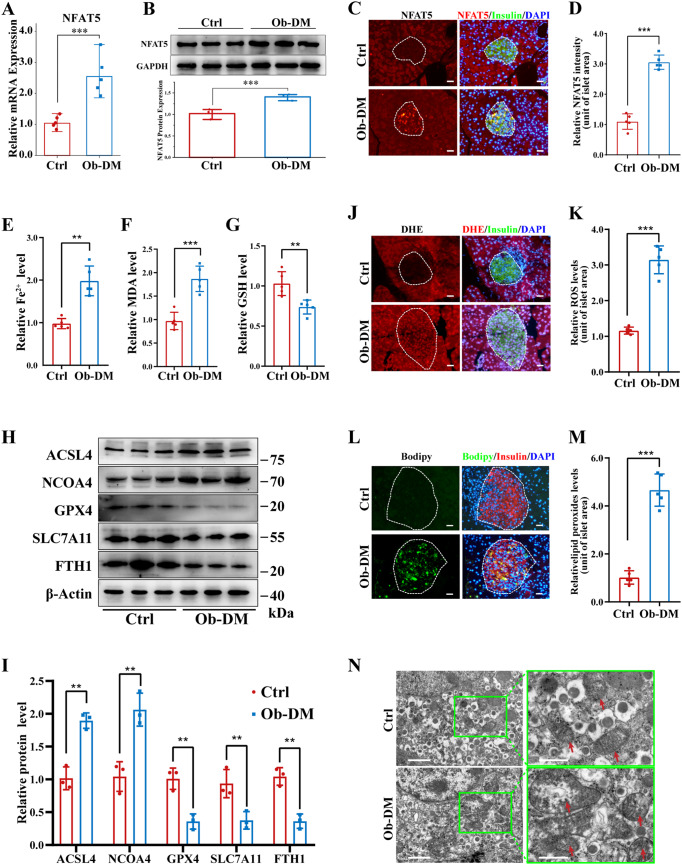
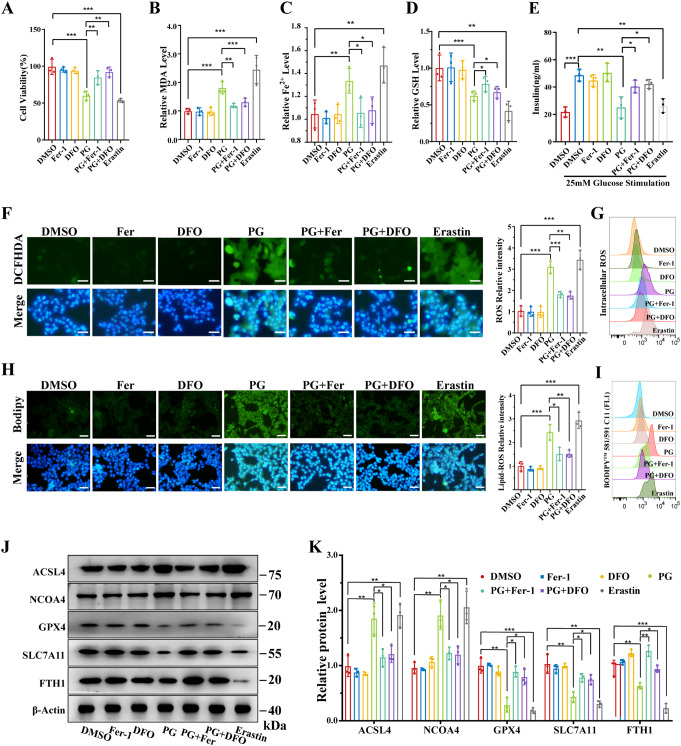
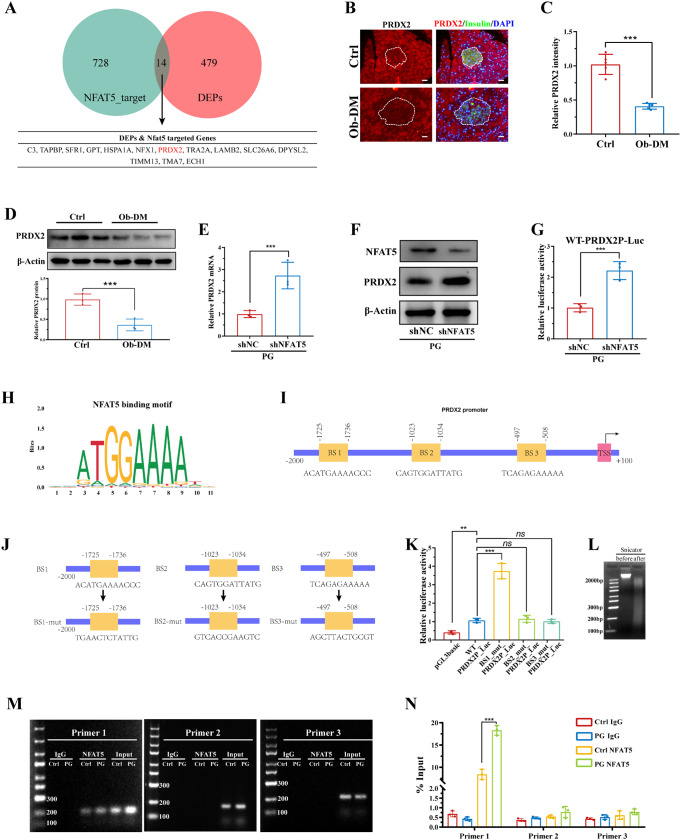
Pancreatic β-cell damage is a critical pathological mechanism in the progression of obese type 2 diabetes mellitus (T2DM). However, the exact underlying mechanism remains unclear. We established an obese T2DM mouse model via high-fat diet feeding. The protein expression profiles of pancreatic tissues from normal and obese T2DM mice were analyzed, revealing that nuclear factor of activated T cells 5 (NFAT5) and ferroptosis are potential mediators and mechanisms of β-cell damage in obese T2DM mice. In vitro, high glucose and palmitate treatment resulted in increased NFAT5 expression and nuclear translocation in MIN6 cells. Inhibition of NFAT5 expression by shRNA significantly reduced ferroptosis and improved the reduction in insulin secretion in palmitic acid and high glucose (PG)-treated MIN6 cells. Luciferase reporter and chromatin immunoprecipitation (ChIP) assays confirmed the ability of NFAT5 to bind to the peroxiredoxin 2 (PRDX2) promoter, leading to the downregulation of PRDX2 transcription. Subsequent rescue experiments confirmed that NFAT5 is involved in PG-induced ferroptosis in MIN6 cells by inhibiting the expression of PRDX2. Finally, we demonstrated that the use of the AAV8-RIP2-miR30-shNFAT5 vector to specifically inhibit the expression of NFAT5 in β-cells significantly diminishes ferroptosis in obese T2DM mice, thereby increasing insulin secretion and improving abnormal glucose tolerance. These findings collectively highlight the therapeutic potential of targeting NFAT5 in β cells to counteract obesity-induced T2DM.
Keywords: Ferroptosis; Nuclear factor of activated T cells 5; Pancreatic β-cells; Peroxiredoxin-2; Type 2 diabetes mellitus.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: The Institutional Animal Care Committee of Central South University approved all the experimental procedures (ethics number CSU-2022-0001-0040). Consent for publication: All the authors agree to publication. Competing interests: The authors declare that they have no conflicts of interest.
Figures





References
-
- Chooi YC, Ding C, Magkos F (2019) The epidemiology of obesity. Metabolism 92:6–10 - PubMed
-
- Fujieda T (2024) Medical Management vs bariatric surgery in type 2 diabetes. JAMA 332(1):76 - PubMed
-
- Kahn SE, Hull RL, Utzschneider KM (2006) Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444(7121):840–846 - PubMed
-
- Prasad MK, Mohandas S, Ramkumar KM (2023) Dysfunctions, molecular mechanisms, and therapeutic strategies of pancreatic β-cells in diabetes. Apoptosis 28(7–8):958–976 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
