

NEAT1 regulates BMSCs aging through disruption of FGF2 nuclear transport
- PMID: 39876006
- PMCID: PMC11776329
- DOI: 10.1186/s13287-025-04156-1
NEAT1 regulates BMSCs aging through disruption of FGF2 nuclear transport
Abstract
Background: The aging of bone marrow mesenchymal stem cells (BMSCs) impairs bone tissue regeneration, contributing to skeletal disorders. LncRNA NEAT1 is considered as a proliferative inhibitory role during cellular senescence, but the relevant mechanisms remain insufficient. This study aims to elucidate how NEAT1 regulates mitotic proteins during BMSCs aging.
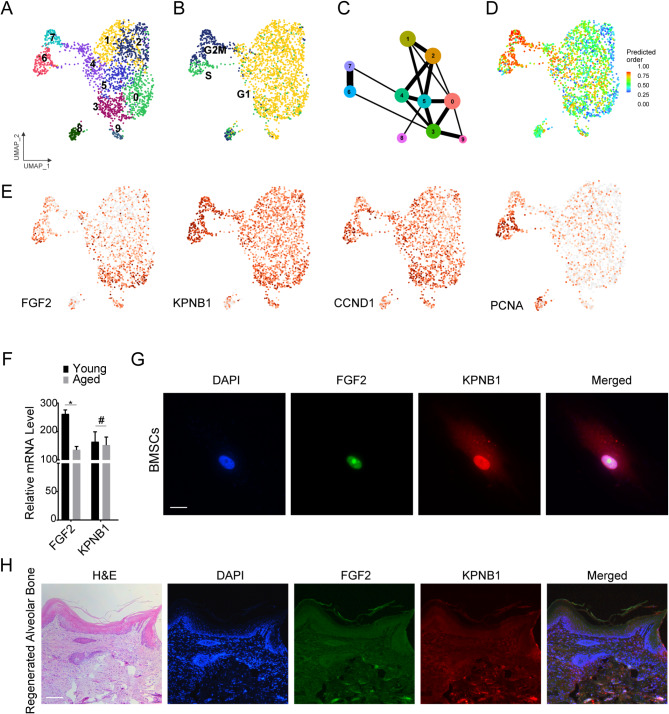
Methods: BMSCs were isolated from alveolar bone of human volunteers aged 26-33 (young) and 66-78 (aged). NEAT1 expression and distribution changes during aging process were observed using fluorescence in situ hybridization (FISH) in young (3 months) and aged (18 months) mice or human BMSCs. Subsequent RNA pulldown and proteomic analyses, along with single-cell analysis, immunofluorescence, RNA immunoprecipitation (RIP), and co-immunoprecipitation (Co-IP), were conducted to investigate that NEAT1 impairs the nuclear transport of mitotic FGF2 and contributes to BMSCs aging.
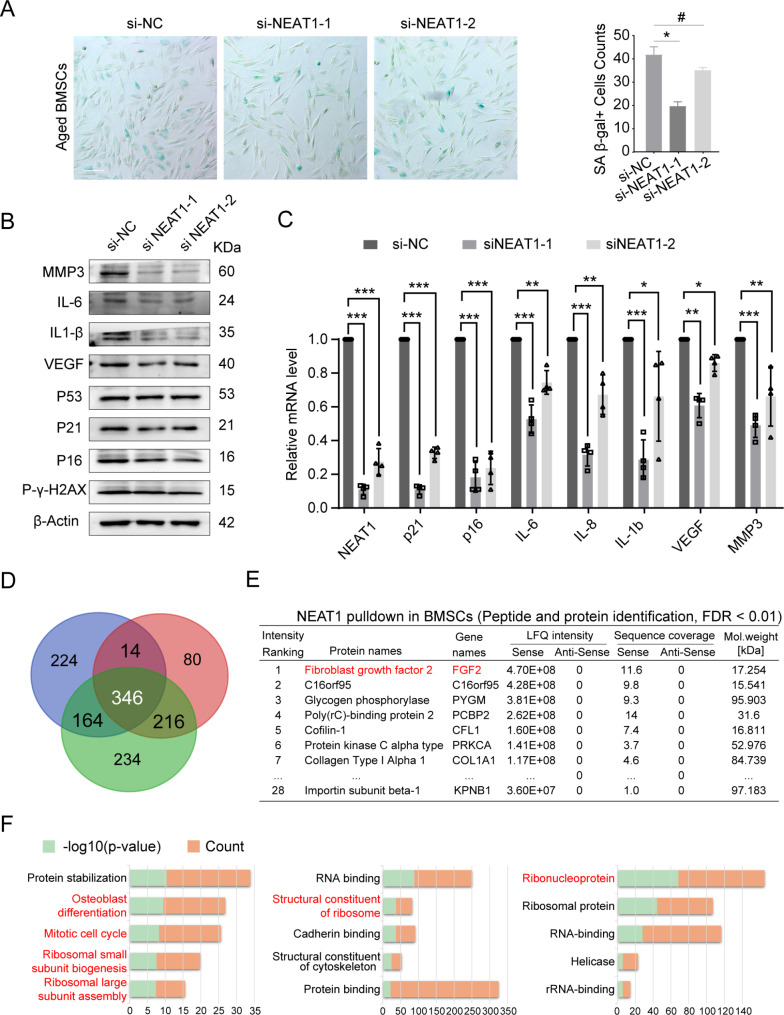
Results: We reveal that NEAT1 undergoes significant upregulated and shifts from nucleus to cytoplasm in bone marrow and BMSCs during aging process. In which, the expression correlates with nuclear DNA content during karyokinesis, suggesting a link to mitogenic factor. Within NEAT1 knockdown, hallmarks of cellular aging, including senescence-associated secretory phenotype (SASP), p16, and p21, were significantly downregulated. RNA pulldown and proteomic analyses further identify NEAT1 involved in osteoblast differentiation, mitotic cell cycle, and ribosome biogenesis, highlighting its role in maintaining BMSCs differentiation and proliferation. Notably, as an essential growth factor of BMSCs, Fibroblast Growth Factor 2 (FGF2) directly abundant binds to NEAT1 and the sites enriched with nuclear localization motifs. Importantly, NEAT1 decreased the interaction between FGF2 and Karyopherin Subunit Beta 1 (KPNB1), influencing the nuclear transport of mitogenic FGF2.
Conclusions: Our findings position NEAT1 as a critical regulator of mitogenic protein networks that govern BMSC aging. Targeting NEAT1 might offer novel therapeutic strategies to rejuvenate aged BMSCs.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval and consent to participate: This study was performed in line with the principles of the Declaration of Helsinki, and written informed consent was obtained from all individual participants before being enrolled in the study. The study design, protocol and informed consent were approved and adopted by the Ethical Committee at Anhui Medical University and the College & Hospital of Stomatology of Anhui Medical University. The project “Collecting jaw bone fragments during dental implant surgery to prepare primary human bone marrow mesenchymal stem cells” was approved on October 20, 2021 with approval number of T2021014. The project “Animal study on the mechanism of aging jaw bone and related BMSCs” was approved on May 30, 2023 with approval number of LLSC20232087. Consent for publication: Not applicable. Competing interests: All authors declare no competing interests. All co-authors have seen and agreed with the contents of the manuscript and there is no financial interest to report.
Figures



Similar articles
-
LncRNA NEAT1 controls the lineage fates of BMSCs during skeletal aging by impairing mitochondrial function and pluripotency maintenance.Cell Death Differ. 2022 Feb;29(2):351-365. doi: 10.1038/s41418-021-00858-0. Epub 2021 Sep 8. Cell Death Differ. 2022. PMID: 34497381 Free PMC article.
-
Fibroblast growth factor 2 enhances BMSC stemness through ITGA2-dependent PI3K/AKT pathway activation.J Cell Physiol. 2024 Dec;239(12):e31423. doi: 10.1002/jcp.31423. Epub 2024 Aug 26. J Cell Physiol. 2024. PMID: 39188080
-
Targeting long noncoding RNA PMIF facilitates osteoprogenitor cells migrating to bone formation surface to promote bone formation during aging.Theranostics. 2021 Mar 20;11(11):5585-5604. doi: 10.7150/thno.54477. eCollection 2021. Theranostics. 2021. PMID: 33859765 Free PMC article.
-
The role of long noncoding RNA (lncRNA) nuclear-enriched abundant transcript 1 (NEAT1) in immune diseases.Transpl Immunol. 2022 Dec;75:101716. doi: 10.1016/j.trim.2022.101716. Epub 2022 Sep 17. Transpl Immunol. 2022. PMID: 36126903 Review.
-
Molecular anatomy of the architectural NEAT1 noncoding RNA: The domains, interactors, and biogenesis pathway required to build phase-separated nuclear paraspeckles.Wiley Interdiscip Rev RNA. 2019 Nov;10(6):e1545. doi: 10.1002/wrna.1545. Epub 2019 May 1. Wiley Interdiscip Rev RNA. 2019. PMID: 31044562 Review.
References
-
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: an expanding universe. Cell. 2023;186(2):243–78. - PubMed
-
- Hou Y, Dan X, Babbar M, Wei Y, Hasselbalch SG, Croteau DL, Bohr VA. Ageing as a risk factor for neurodegenerative disease. Nat Rev Neurol. 2019;15(10):565–81. - PubMed
MeSH terms
Substances
Grants and funding
- 82201026/National Natural Science Foundation of China
- 2408085MH221/Natural Science Foundation of Anhui Province
- YQYB2024013/Outstanding Young Teacher Development Program for Higher Education Institutions of Anhui Province
- 2023xkj041/The School Foundation of Anhui Medical University
- S202410366087/Anhui Medical University Student Innovation and Entrepreneurship Training Program
LinkOut - more resources
Full Text Sources
Miscellaneous
