

Allogeneic DNT cell therapy synergizes with T cells to promote anti-leukemic activities while suppressing GvHD
- PMID: 39876025
- PMCID: PMC11773727
- DOI: 10.1186/s13046-024-03247-w
Allogeneic DNT cell therapy synergizes with T cells to promote anti-leukemic activities while suppressing GvHD
Abstract
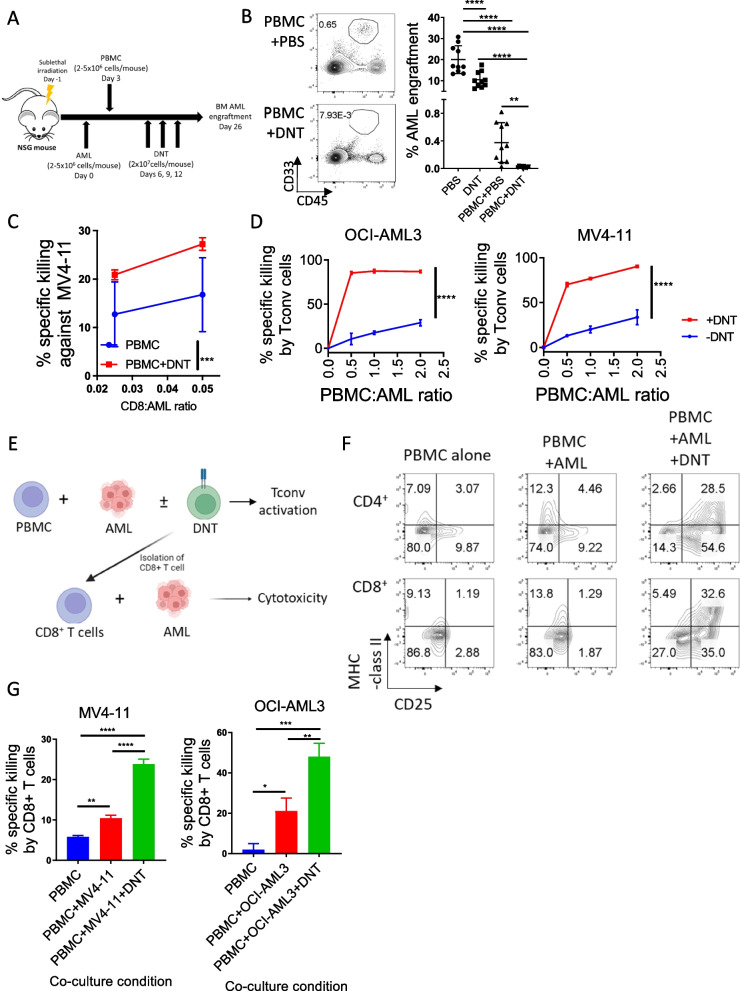
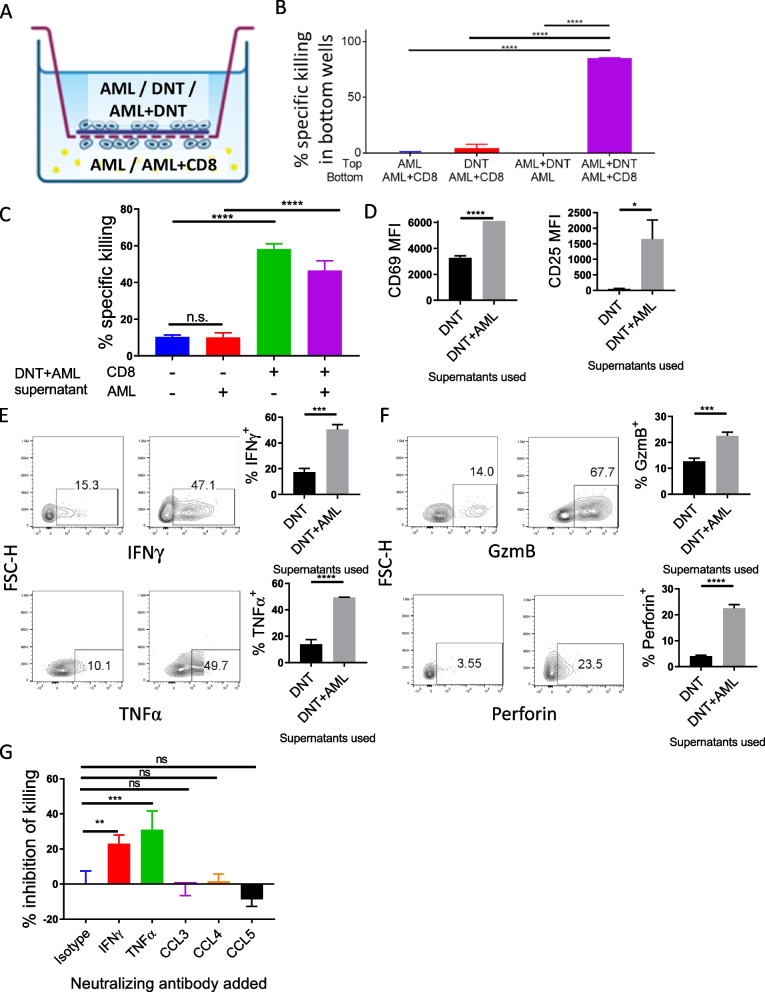
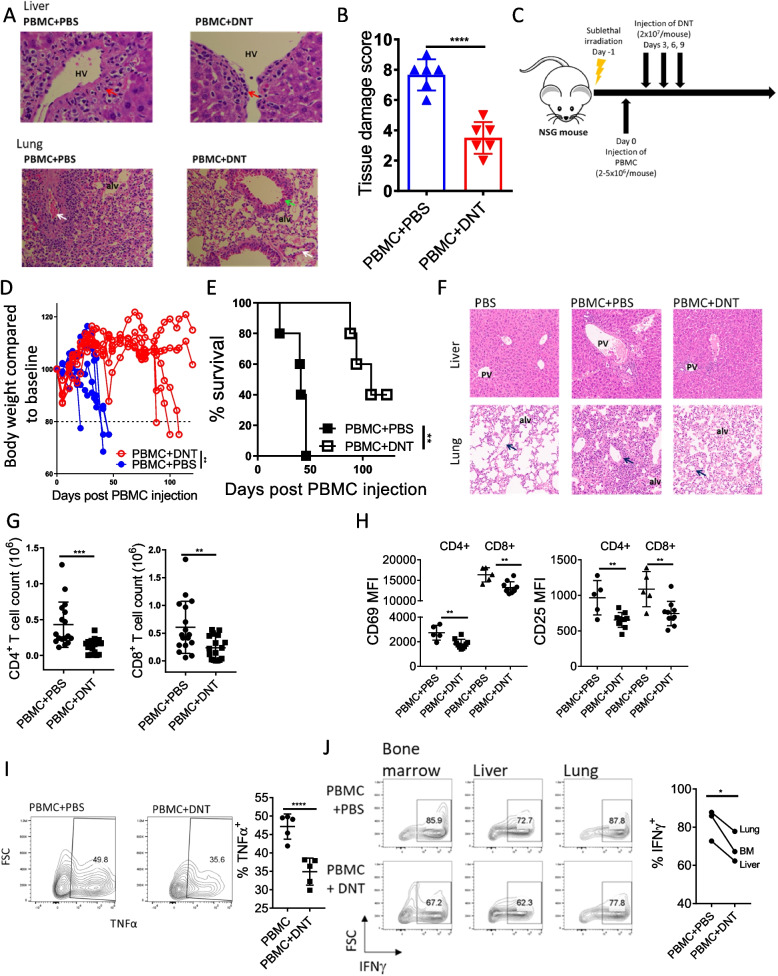
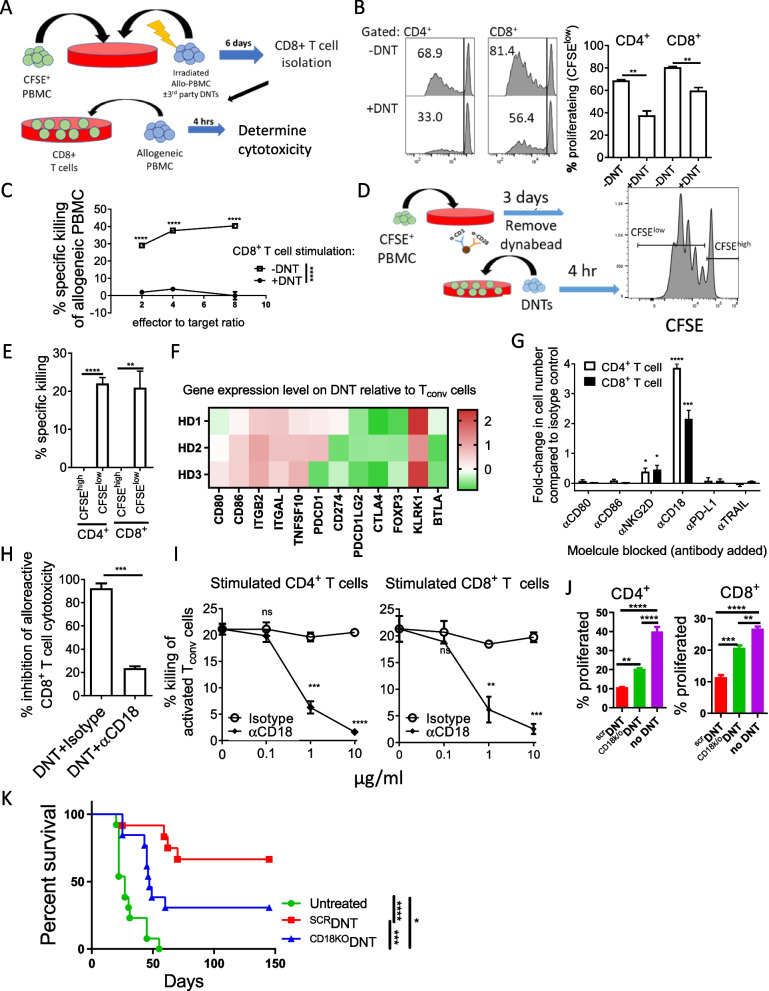
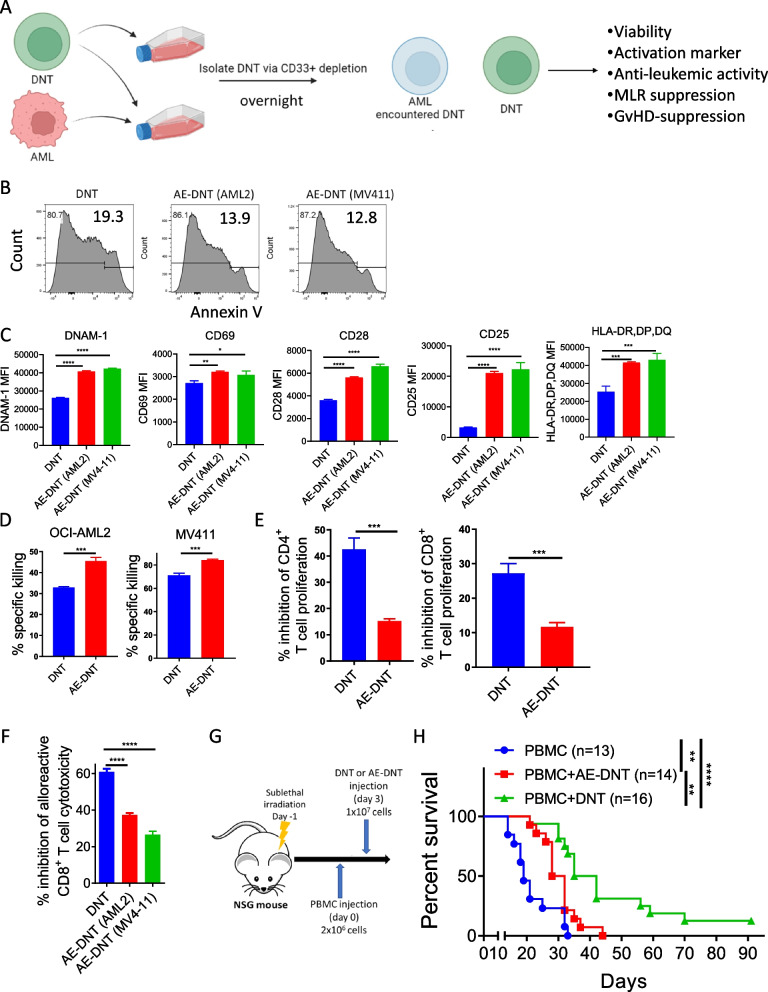
Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is a second-line treatment with curative potential for leukemia patients. However, the prognosis of allo-HSCT patients with disease relapse or graft-versus-host disease (GvHD) is poor. CD4+ or CD8+ conventional T (Tconv) cells are critically involved in mediating anti-leukemic immune responses to prevent relapse and detrimental GvHD. Hence, treatment for one increases the risk of the other. Thus, therapeutic strategies that can address relapse and GvHD are considered the Holy Grail of allo-HSCT. CD3+CD4-CD8- double-negative T cells (DNTs) are unconventional mature T cells with potent anti-leukemia effects with "off-the-shelf" potential. A phase I clinical trial demonstrated the feasibility, safety, and potential efficacy of allogeneic DNT therapy for patients with relapsing acute myeloid leukemia (AML) post-allo-HSCT. Here, we studied the impact of DNTs on the anti-leukemic and GvHD-inducing activities of Tconv cells. DNTs synergized with Tconv cells to mediate superior anti-leukemic activity. Mechanistically, DNTs released soluble factors which activated and evoked potent anti-leukemic activities of Tconv cells. In contrast, DNTs suppressed GvHD-inducing activities of Tconv cells in a CD18-dependent manner by mediating cytotoxicity against proliferative Tconv cells. The seemingly opposite immunological activities of DNTs were dictated by the presence or absence of AML cells. Collectively, these results support the potential of DNTs as an adjuvant to allo-HSCT to address both disease relapse and GvHD.
Keywords: Allogeneic hematopoietic stem cell transplantation; Donor lymphocyte infusion; Double negative T cell; Graft-versus-host disease; Graft-versus-leukemia.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Consent for publication: All of the authors concur with the submission of this manuscript. Competing interests: L.Z. has financial interests (e.g., holdings/shares) in WYZE Biotech Co Ltd and previously received research funding and consulting fee/honorarium from the Company. L.Z., J.B.L and H.K. are inventors of several patents related to DNT cell technology, and L.Z. received license fees. The remaining authors declare no conflict of interest.
Figures
References
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
