

Prior vaccination prevents overactivation of innate immune responses during COVID-19 breakthrough infection
- PMID: 39879318
- PMCID: PMC12142493
- DOI: 10.1126/scitranslmed.adq1086
Prior vaccination prevents overactivation of innate immune responses during COVID-19 breakthrough infection
Abstract
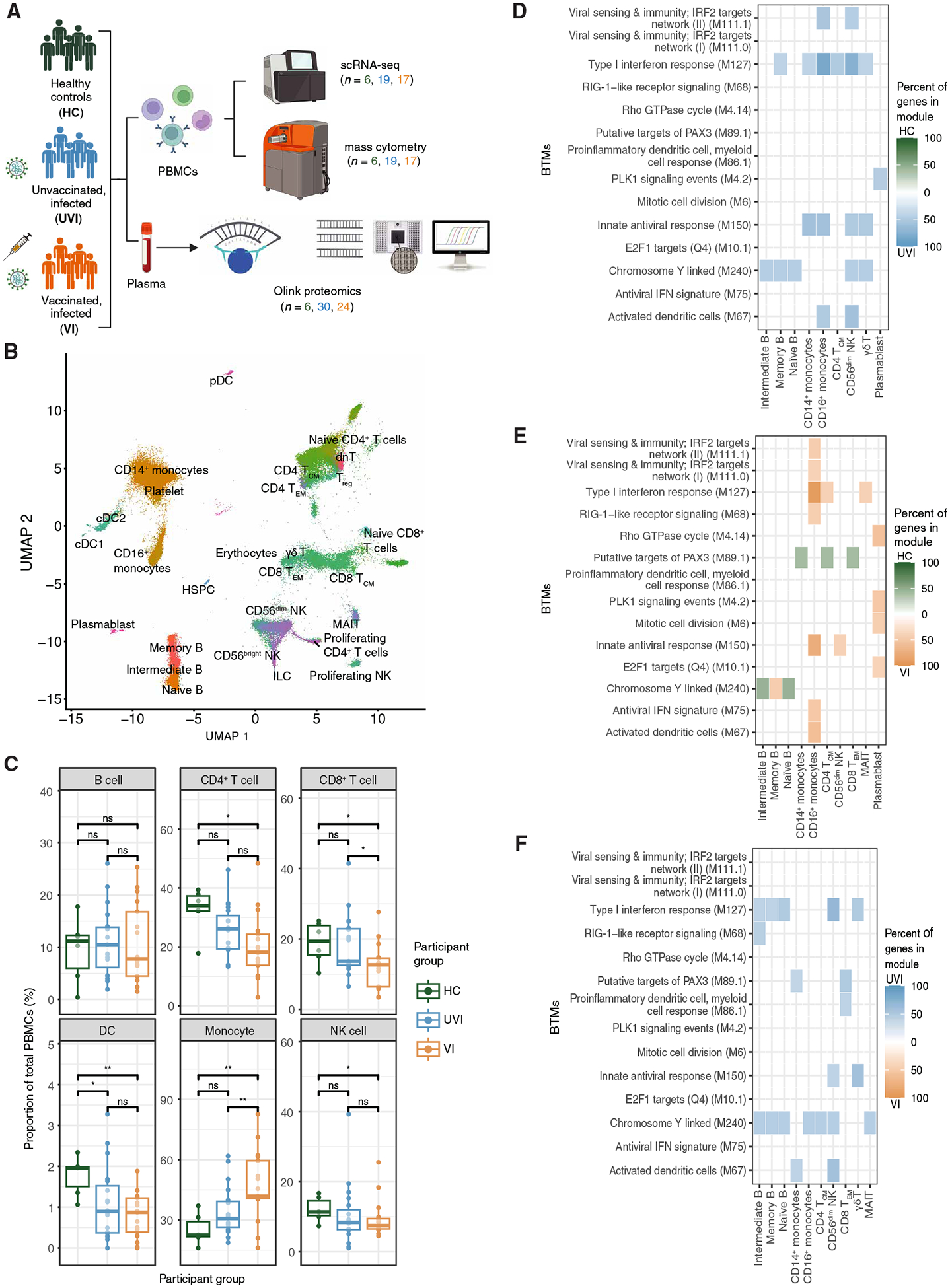
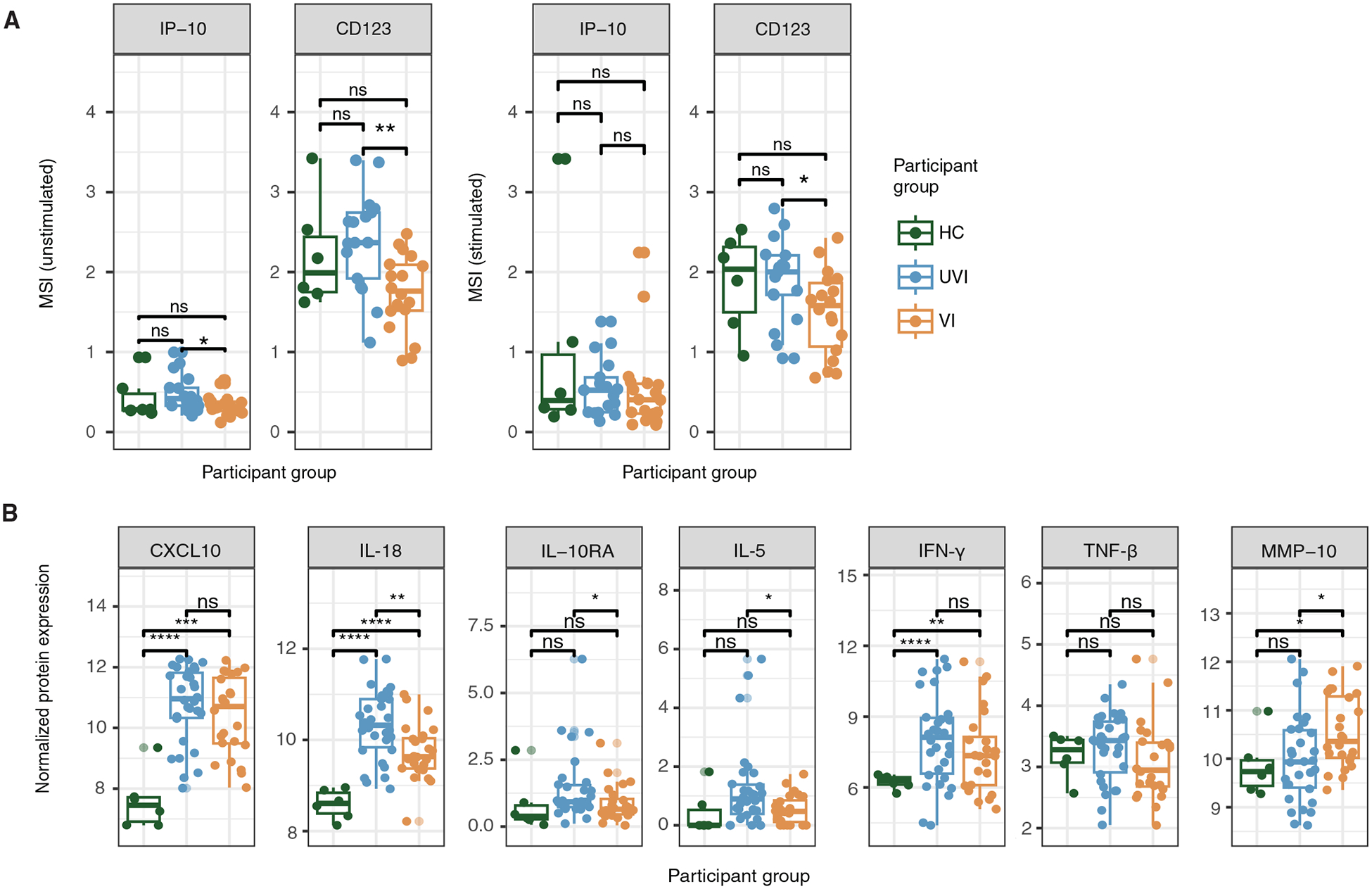
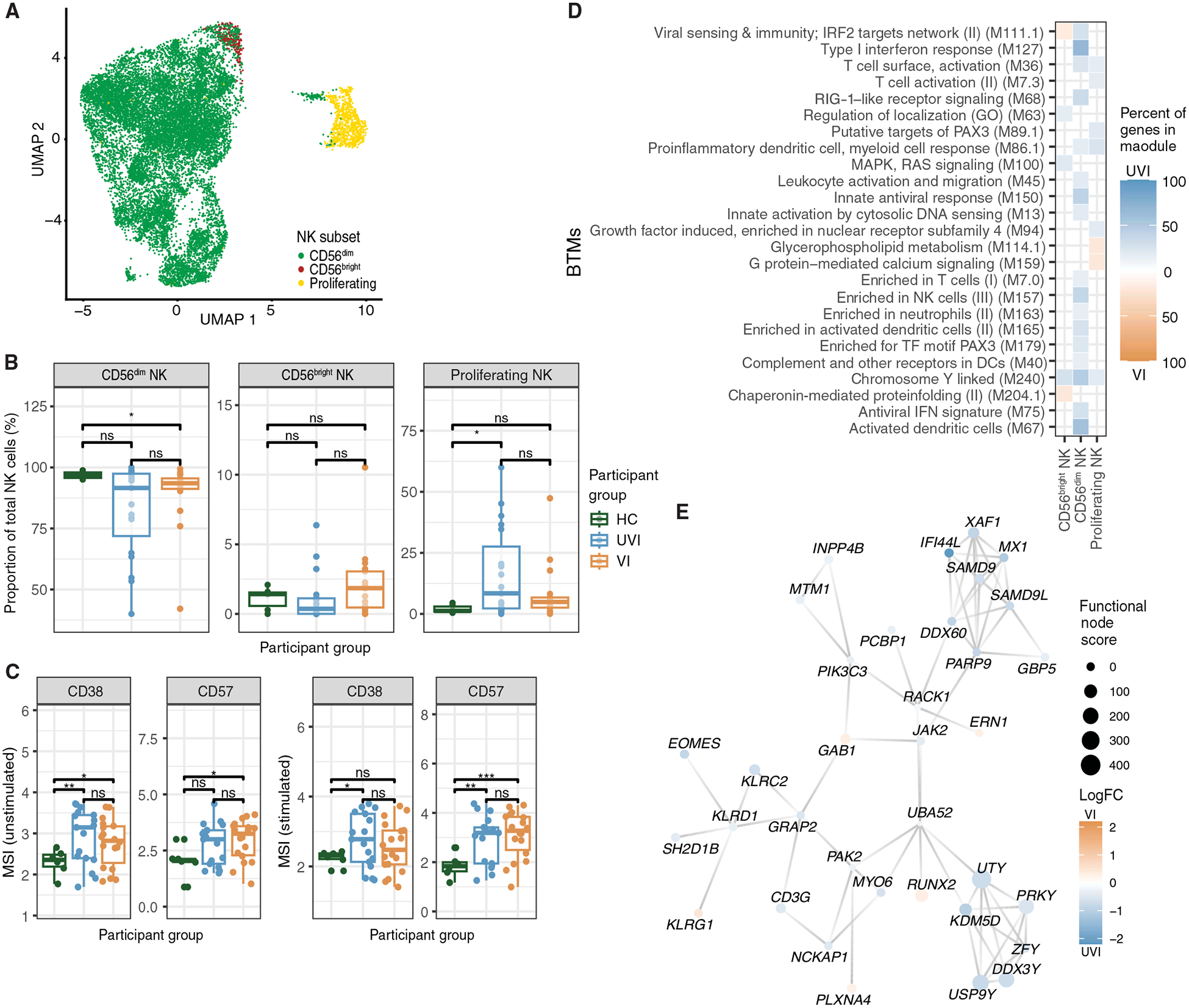
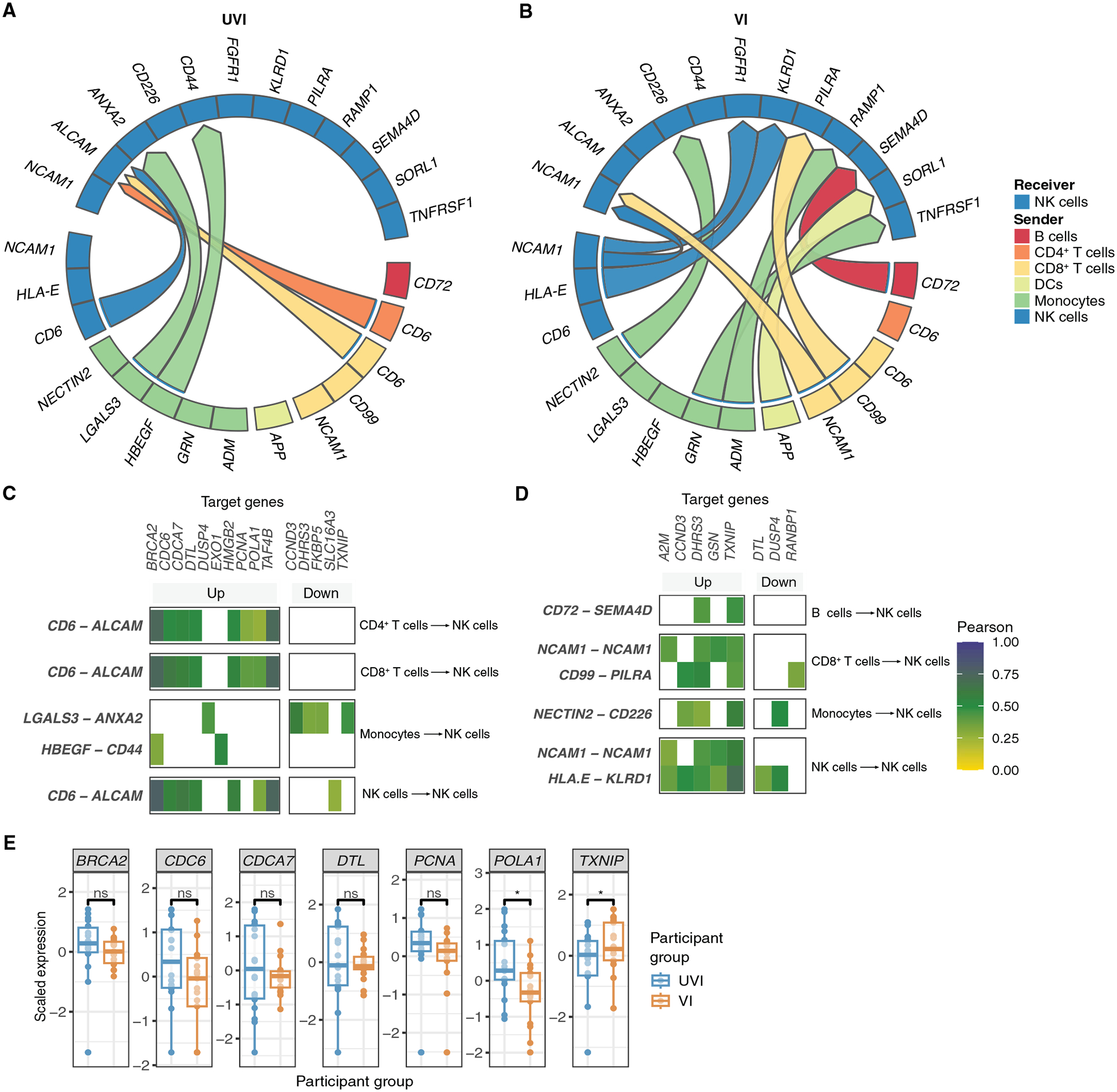
At this stage in the COVID-19 pandemic, most infections are "breakthrough" infections that occur in individuals with prior severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) exposure. To refine long-term vaccine strategies against emerging variants, we examined both innate and adaptive immunity in breakthrough infections. We performed single-cell transcriptomic, proteomic, and functional profiling of primary and breakthrough infections to compare immune responses from unvaccinated and vaccinated individuals during the SARS-CoV-2 Delta wave. Breakthrough infections were characterized by a less activated transcriptomic profile in monocytes and natural killer cells, with induction of pathways limiting monocyte migratory potential and natural killer cell proliferation. Furthermore, we observed a female-specific increase in transcriptomic and proteomic activation of multiple innate immune cell subsets during breakthrough infections. These insights suggest that prior SARS-CoV-2 vaccination prevents overactivation of innate immune responses during breakthrough infections with discernible sex-specific patterns and underscore the potential of harnessing vaccines in mitigating pathologic immune responses resulting from overactivation.
Conflict of interest statement
Figures




References
-
- Fan Q, Shi J, Yang Y, Tang G, Jiang M, Li J, Tang J, Li L, Wen X, Zhang L, Deng X, Wang Y, Lan Y, Li L, Peng P, Tong Y, Lu H, Yan L, Liu Y, Cai S, Li Y, Mo X, Li M, Deng X, Hu Z, Yu H, Hu F, Liu J, Tang X, Li F, Clinical characteristics and immune profile alterations in vaccinated individuals with breakthrough Delta SARS-CoV-2 infections. Nat. Commun 13, 3979 (2022). - PMC - PubMed
-
- Collier A-RY, Brown CM, Mcmahan K, Yu J, Liu J, Jacob-Dolan C, Chandrashekar A, Tierney D, Ansel JL, Rowe M, Sellers D, Ahmad K, Aguayo R, Anioke T, Gardner S, Siamatu M, Bermudez Rivera L, Hacker MR, Madoff LC, Barouch DH, Immune responses in fully vaccinated individuals following breakthrough infection with the SARS-CoV-2 Delta variant in Provincetown, Massachusetts. medRxiv 21265113 [Preprint] (2021). 10.1101/2021.10.18.21265113. - DOI - PMC - PubMed
-
- Koutsakos M, Lee WS, Reynaldi A, Tan H-X, Gare G, Kinsella P, Liew KC, Taiaroa G, Williamson DA, Kent HE, Stadler E, Cromer D, Khoury DS, Wheatley AK, Juno JA, Davenport MP, Kent SJ, The magnitude and timing of recalled immunity after breakthrough infection is shaped by SARS-CoV-2 variants. Immunity 55, 1316–1326.e4 (2022). - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
