

Sucrose-preferring gut microbes prevent host obesity by producing exopolysaccharides
- PMID: 39880823
- PMCID: PMC11779931
- DOI: 10.1038/s41467-025-56470-0
Sucrose-preferring gut microbes prevent host obesity by producing exopolysaccharides
Abstract
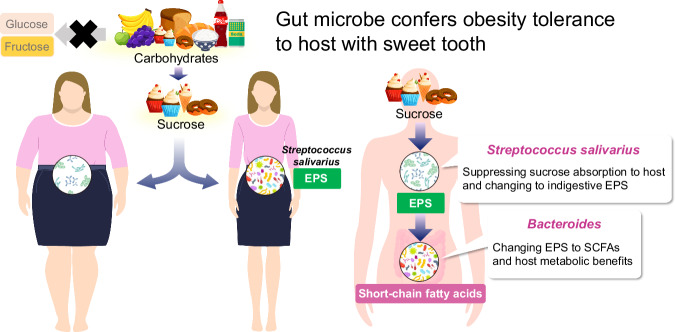
Commensal bacteria affect host health by producing various metabolites from dietary carbohydrates via bacterial glycometabolism; however, the underlying mechanism of action remains unclear. Here, we identified Streptococcus salivarius as a unique anti-obesity commensal bacterium. We found that S. salivarius may prevent host obesity caused by excess sucrose intake via the exopolysaccharide (EPS) -short-chain fatty acid (SCFA) -carbohydrate metabolic axis in male mice. Healthy human donor-derived S. salivarius produced high EPS levels from sucrose but not from other sugars. S. salivarius abundance was significantly decreased in human donors with obesity compared with that in healthy donors, and the EPS-SCFA bacterial carbohydrate metabolic process was attenuated. Our findings reveal an important mechanism by which host-commensal interactions in glycometabolism affect energy regulation, suggesting an approach for preventing lifestyle-related diseases via prebiotics and probiotics by targeting bacteria and EPS metabolites.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: H.S., K.H., and D.S. are employees of Noster Inc. The other authors declare no competing interests.
Figures






References
-
- Kahn, S. E., Hull, R. L. & Utzschneider, K. M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature444, 840–846 (2006). - PubMed
-
- Zimmet, P., Alberti, K. G. & Shaw, J. Global and societal implications of the diabetes epidemic. Nature414, 782–787 (2001). - PubMed
-
- Cori, C. F. Mammalian carbohydrate metabolism. Physiol. Rev.11, 144–275 (1931).
-
- Tappy, L. & Lê, K. A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev.90, 23–46 (2010). - PubMed
-
- Flint, H. J., Bayer, E. A., Rincon, M. T., Lamed, R. & White, B. A. Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat. Rev. Microbiol.6, 121–131 (2008). - PubMed
MeSH terms
Substances
Grants and funding
- JP17gm1010007/Japan Agency for Medical Research and Development (AMED)
- JP23gm1510011/Japan Agency for Medical Research and Development (AMED)
- JP21H04862/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP22K17771/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP21K19813/MEXT | Japan Society for the Promotion of Science (JSPS)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
