

Copper-dependent halogenase catalyses unactivated C-H bond functionalization
- PMID: 39880944
- PMCID: PMC12145154
- DOI: 10.1038/s41586-024-08362-4
Copper-dependent halogenase catalyses unactivated C-H bond functionalization
Abstract
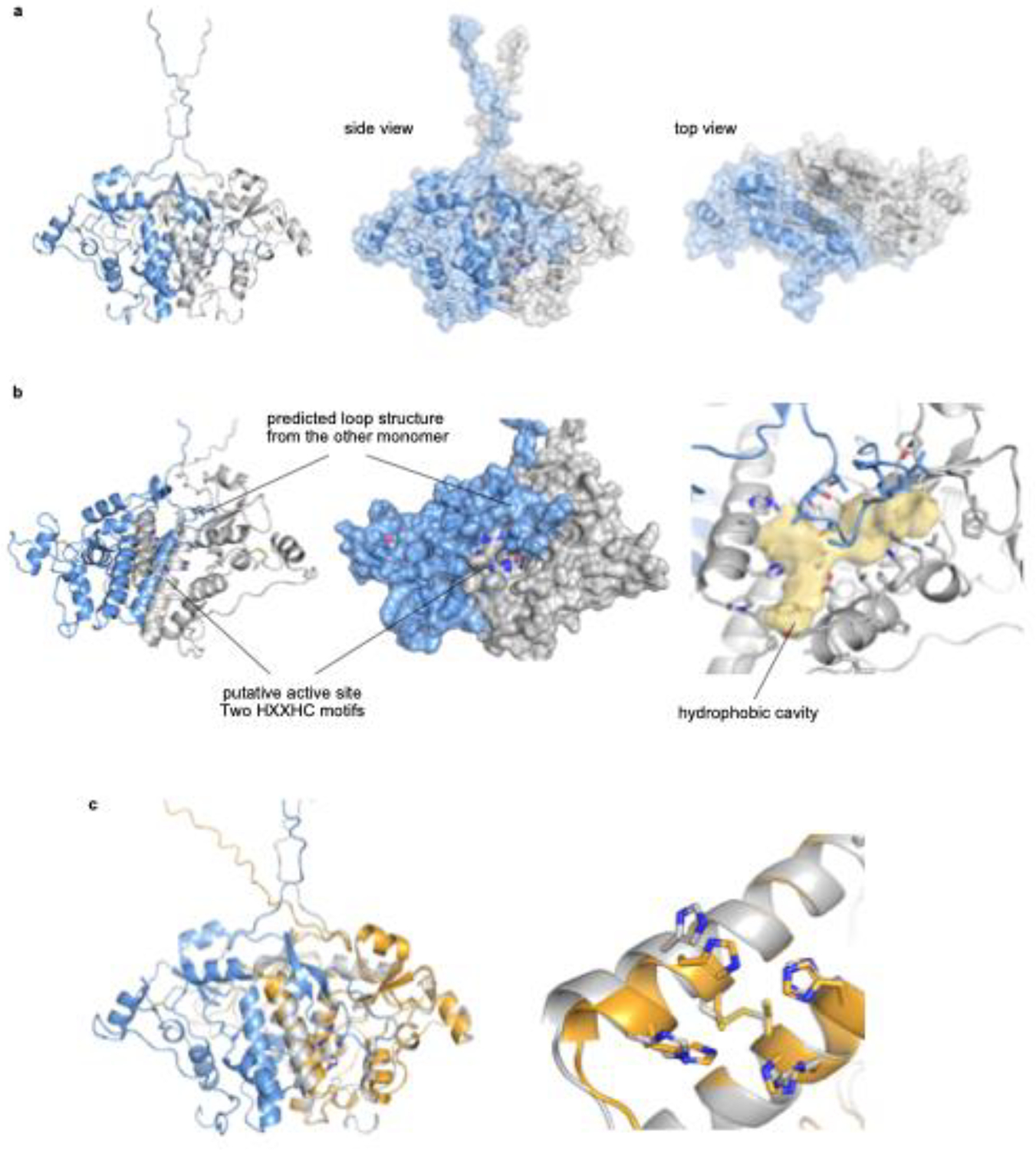
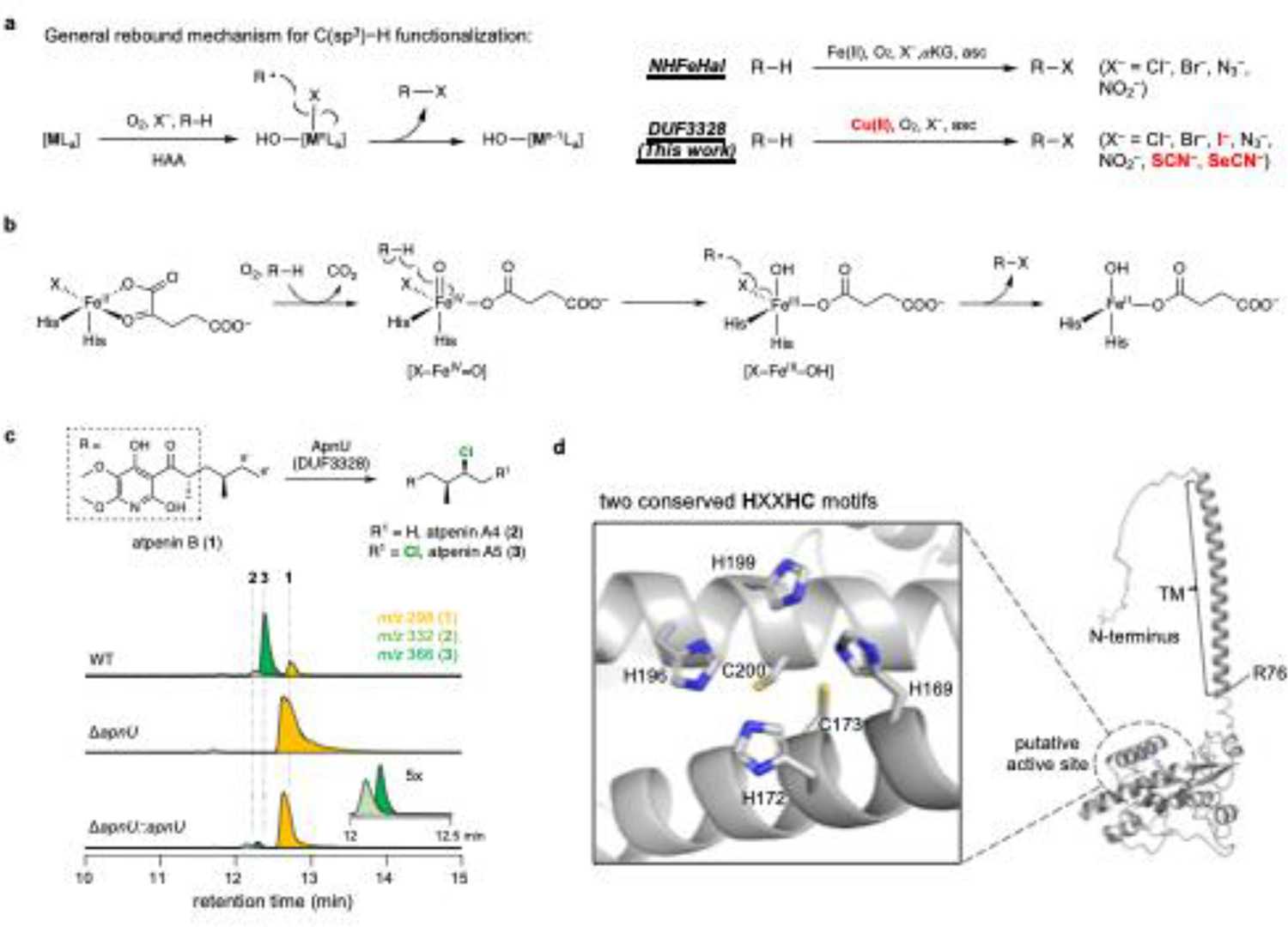
Carbon-hydrogen (C-H) bonds are the foundation of essentially every organic molecule, making them an ideal place to do chemical synthesis. The key challenge is achieving selectivity for one particular C(sp3)-H bond1-3. In recent years, metalloenzymes have been found to perform C(sp3)-H bond functionalization4,5. Despite substantial progresses in the past two decades6,7, enzymatic halogenation and pseudohalogenation of unactivated C(sp3)-H-providing a functional handle for further modification-have been achieved with only non-haem iron/α-ketoglutarate-dependent halogenases, and are therefore limited by the chemistry possible with these enzymes8. Here we report the discovery and characterization of a previously unknown halogenase ApnU, part of a protein family containing domain of unknown function 3328 (DUF3328). ApnU uses copper in its active site to catalyse iterative chlorinations on multiple unactivated C(sp3)-H bonds. By taking advantage of the softer copper centre, we demonstrate that ApnU can catalyse unprecedented enzymatic C(sp3)-H bond functionalization such as iodination and thiocyanation. Using biochemical characterization and proteomics analysis, we identified the functional oligomeric state of ApnU as a covalently linked homodimer, which contains three essential pairs-one interchain and two intrachain-of disulfide bonds. The metal-coordination active site in ApnU consists of binuclear type II copper centres, as revealed by electron paramagnetic resonance spectroscopy. This discovery expands the enzymatic capability of C(sp3)-H halogenases and provides a foundational understanding of this family of binuclear copper-dependent oxidative enzymes.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures












Similar articles
-
Biochemical Studies of a Cyanobacterial Halogenase Support the Involvement of a Dimetal Cofactor.Biochemistry. 2025 May 20;64(10):2173-2180. doi: 10.1021/acs.biochem.4c00720. Epub 2025 Apr 29. Biochemistry. 2025. PMID: 40300765 Free PMC article.
-
Light-Driven C(sp3)-C(sp3) Bond Functionalizations Enabled by the PCET Activation of Alcohol O-H Bonds.Acc Chem Res. 2025 Jul 1;58(13):2061-2071. doi: 10.1021/acs.accounts.5c00246. Epub 2025 Jun 13. Acc Chem Res. 2025. PMID: 40511721
-
Copper-Dependent Hydroxylation Catalyzed by the DUF3328 Enzyme CctR.Org Lett. 2025 Aug 1;27(30):8178-8182. doi: 10.1021/acs.orglett.5c02280. Epub 2025 Jul 22. Org Lett. 2025. PMID: 40693726
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Transition-Metal-Catalyzed C(sp3)-H Bond Activation through Coordination-Assisted and Radical Mechanisms: Recent Advances and Mechanistic Insight.Top Curr Chem (Cham). 2025 Aug 26;383(3):33. doi: 10.1007/s41061-025-00511-1. Top Curr Chem (Cham). 2025. PMID: 40856896 Review.
Cited by
-
Fungal RiPPs Side Chain Macrocyclization Catalyzed by Copper-Dependent DUF3328 Enzyme.J Am Chem Soc. 2025 Mar 12;147(10):8113-8117. doi: 10.1021/jacs.4c18770. Epub 2025 Mar 3. J Am Chem Soc. 2025. PMID: 40029812
-
Peptide Recognition Sequence Guides Catalytic Side Chain Cross-Linking of Plant Peptides by Copper-Dependent Cyclases.J Am Chem Soc. 2025 Jun 18;147(24):20284-20293. doi: 10.1021/jacs.4c15470. Epub 2025 Jun 5. J Am Chem Soc. 2025. PMID: 40470882 Free PMC article.
-
SeqForge: A scalable platform for alignment-based searches, motif detection, and sequence curation across meta/genomic datasets.bioRxiv [Preprint]. 2025 Aug 15:2025.08.12.669971. doi: 10.1101/2025.08.12.669971. bioRxiv. 2025. PMID: 40832300 Free PMC article. Preprint.
-
Biochemical Studies of a Cyanobacterial Halogenase Support the Involvement of a Dimetal Cofactor.Biochemistry. 2025 May 20;64(10):2173-2180. doi: 10.1021/acs.biochem.4c00720. Epub 2025 Apr 29. Biochemistry. 2025. PMID: 40300765 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
