

C-terminal amides mark proteins for degradation via SCF-FBXO31
- PMID: 39880951
- PMCID: PMC11821526
- DOI: 10.1038/s41586-024-08475-w
C-terminal amides mark proteins for degradation via SCF-FBXO31
Abstract
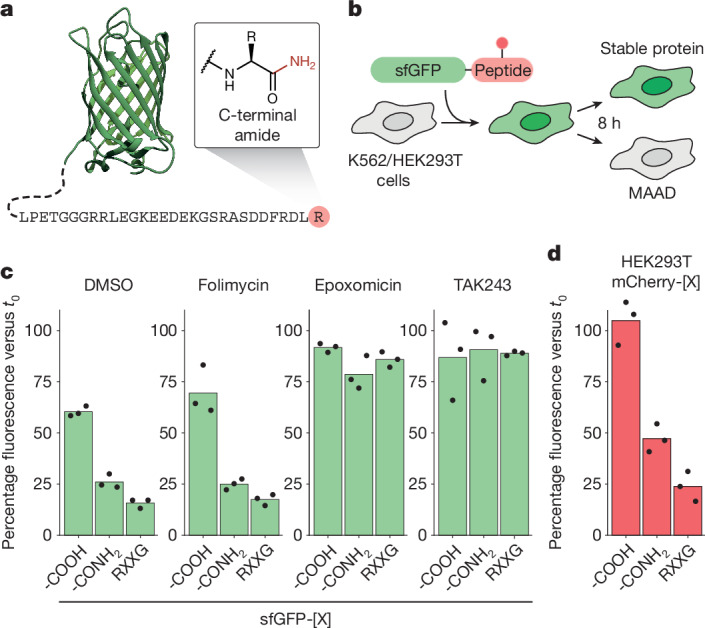
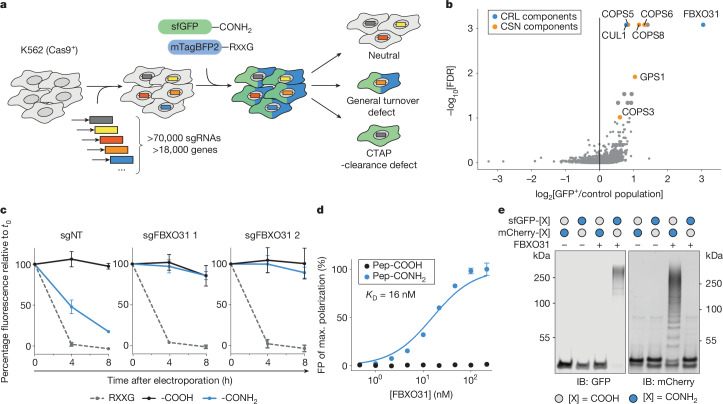
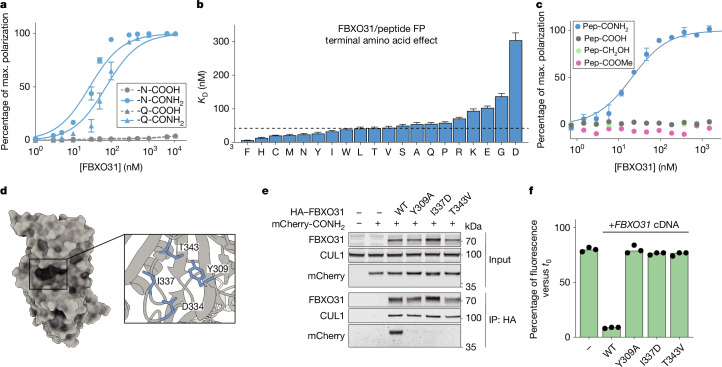
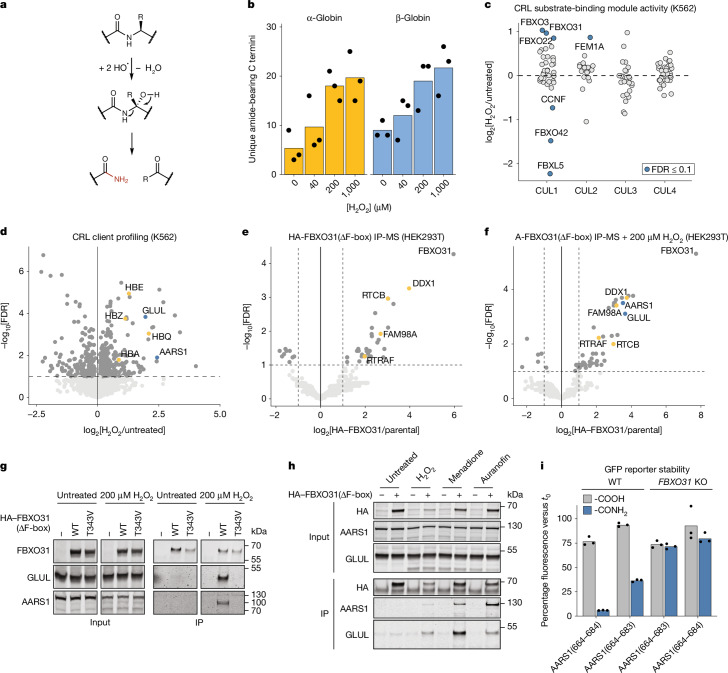
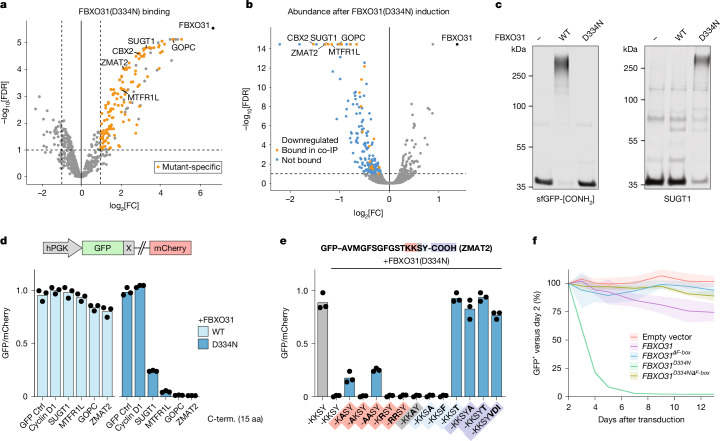
During normal cellular homeostasis, unfolded and mislocalized proteins are recognized and removed, preventing the build-up of toxic byproducts1. When protein homeostasis is perturbed during ageing, neurodegeneration or cellular stress, proteins can accumulate several forms of chemical damage through reactive metabolites2,3. Such modifications have been proposed to trigger the selective removal of chemically marked proteins3-6; however, identifying modifications that are sufficient to induce protein degradation has remained challenging. Here, using a semi-synthetic chemical biology approach coupled to cellular assays, we found that C-terminal amide-bearing proteins (CTAPs) are rapidly cleared from human cells. A CRISPR screen identified FBXO31 as a reader of C-terminal amides. FBXO31 is a substrate receptor for the SKP1-CUL1-F-box protein (SCF) ubiquitin ligase SCF-FBXO31, which ubiquitylates CTAPs for subsequent proteasomal degradation. A conserved binding pocket enables FBXO31 to bind to almost any C-terminal peptide bearing an amide while retaining exquisite selectivity over non-modified clients. This mechanism facilitates binding and turnover of endogenous CTAPs that are formed after oxidative stress. A dominant human mutation found in neurodevelopmental disorders reverses CTAP recognition, such that non-amidated neosubstrates are now degraded and FBXO31 becomes markedly toxic. We propose that CTAPs may represent the vanguard of a largely unexplored class of modified amino acid degrons that could provide a general strategy for selective yet broad surveillance of chemically damaged proteins.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: M.F.M., J.F., R.H., J.W.B. and J.E.C. have filed patent applications related to MAAD discovery and mechanisms of FBXO31 substrate recognition performed in this study (WO/2024/115740, WO/2024/115746; both pending). J.W.B. and J.E.C. are founders of and M.F.M. and J.F. provide consultancy to Serac Biosciences. J.E.C. serves on the scientific advisory board of Mission Therapeutics. B.A.S. serves on the scientific advisory boards of Biotheryx and Proxygen. M.M. is an indirect investor of the Evosep company. The other authors declare no competing interests.
Figures










References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
