

Prominosomes - a particular class of extracellular vesicles containing prominin-1/CD133?
- PMID: 39881297
- PMCID: PMC11776279
- DOI: 10.1186/s12951-025-03102-w
Prominosomes - a particular class of extracellular vesicles containing prominin-1/CD133?
Abstract
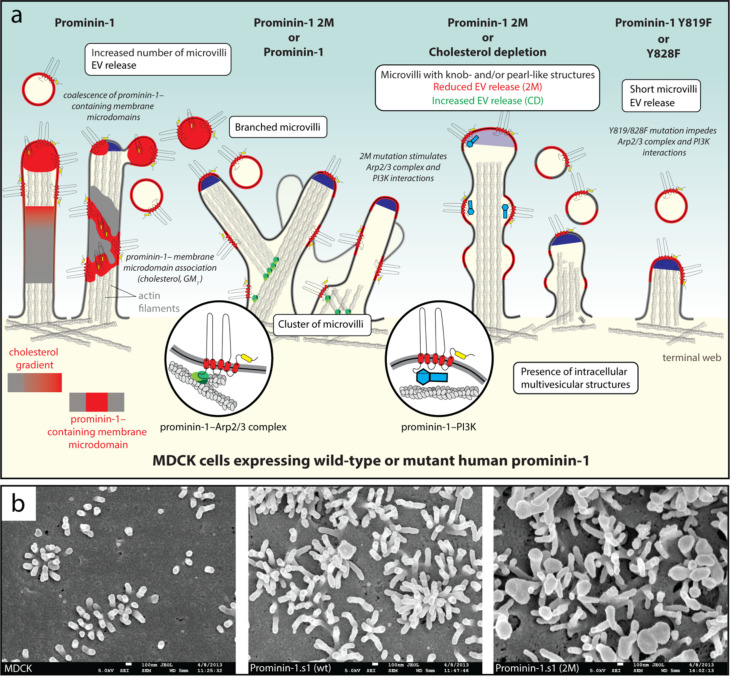
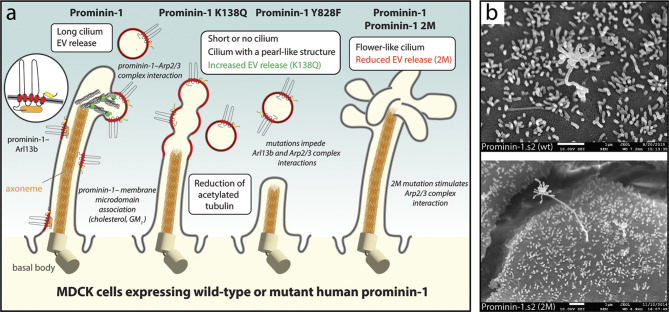
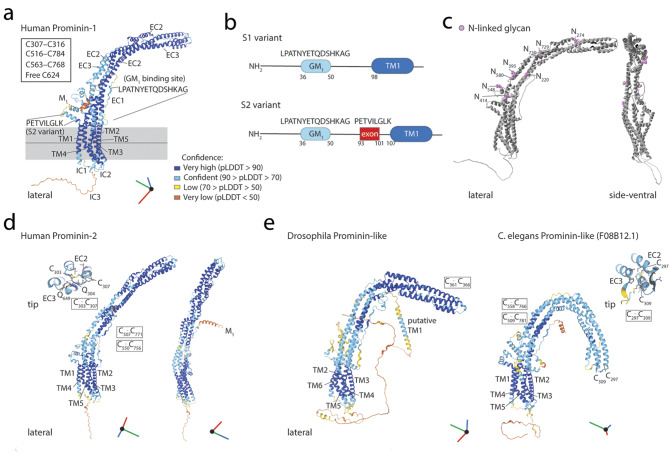
Extracellular membrane vesicles (EVs) offer promising values in various medical fields, e.g., as biomarkers in liquid biopsies or as native (or bioengineered) biological nanocarriers in tissue engineering, regenerative medicine and cancer therapy. Based on their cellular origin EVs can vary considerably in composition and diameter. Cell biological studies on mammalian prominin-1, a cholesterol-binding membrane glycoprotein, have helped to reveal new donor membranes as sources of EVs. For instance, small EVs can originate from microvilli and primary cilia, while large EVs might be produced by transient structures such as retracting cellular extremities of cancer cells during the mitotic rounding process, and the midbody at the end of cytokinesis. Here, we will highlight the various subcellular origins of prominin-1+ EVs, also called prominosomes, and the potential mechanism(s) regulating their formation. We will further discuss the molecular and cellular characteristics of prominin-1, notably those that have a direct effect on the release of prominin-1+ EVs, a process that might be directly implicated in donor cell reprogramming of stem and cancer stem cells. Prominin-1+ EVs also mediate intercellular communication during embryonic development and adult homeostasis in healthy individuals, while disseminating biological information during diseases.
Keywords: CD133; Cell signaling; Cilium; Ectosome; Exosome; Intercellular communication; Lipid droplet; Microvillus; Midbody; Stem cell.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
