

Activation of the conserved Hippo kinases by inflammasome-triggered proteolytic cleavage controls programmed cell death in macrophages
- PMID: 39883842
- PMCID: PMC11804562
- DOI: 10.1073/pnas.2418613122
Activation of the conserved Hippo kinases by inflammasome-triggered proteolytic cleavage controls programmed cell death in macrophages
Abstract
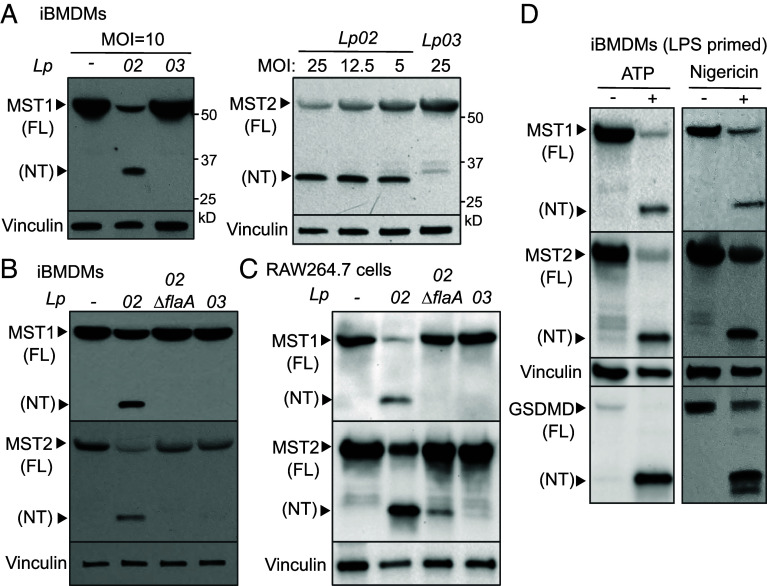
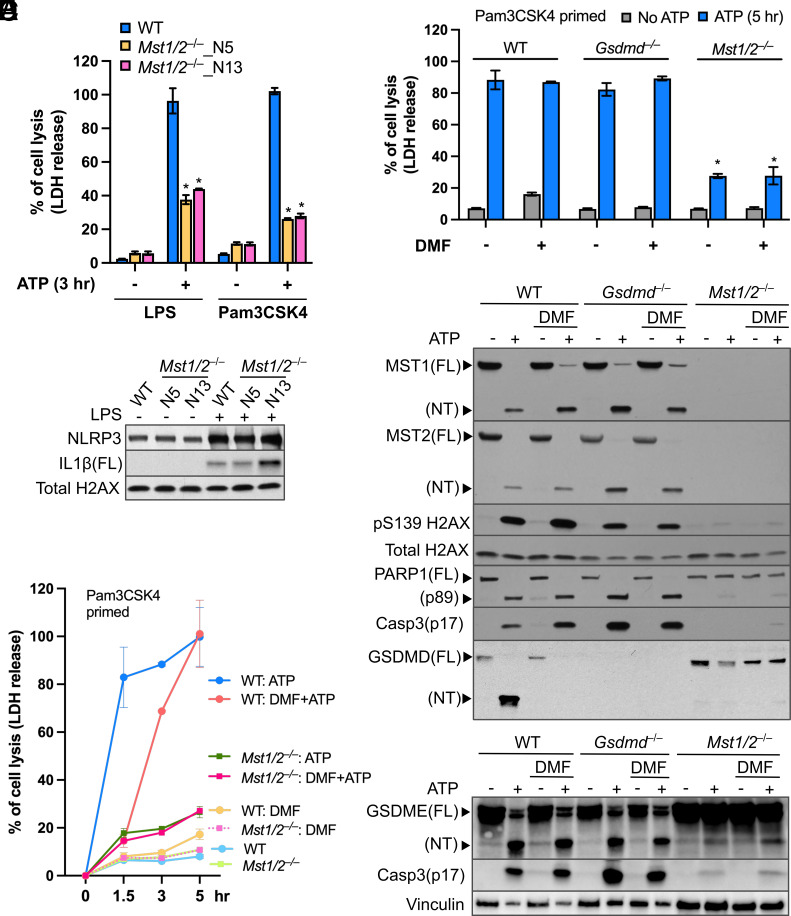
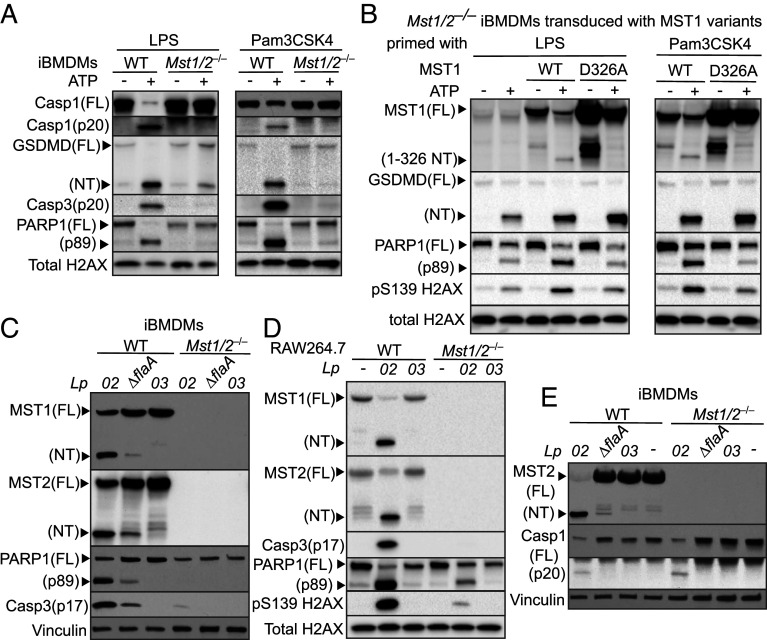
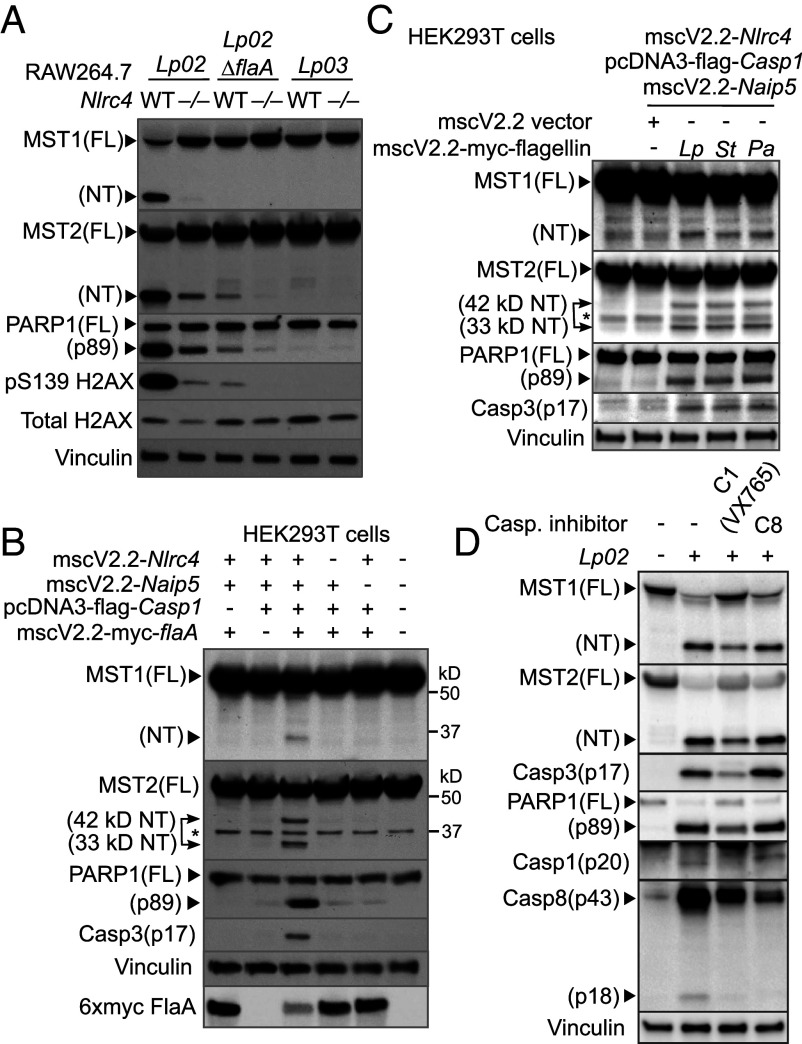
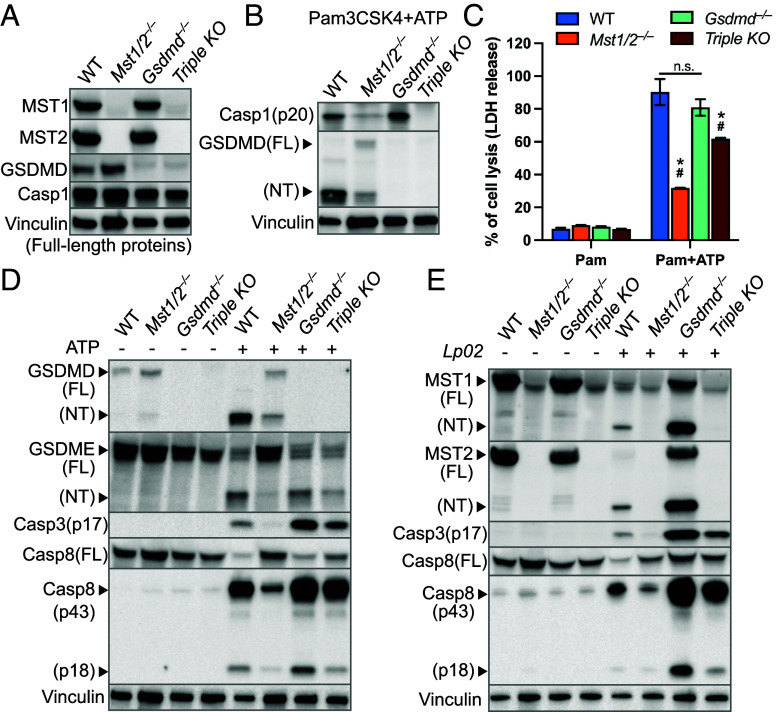
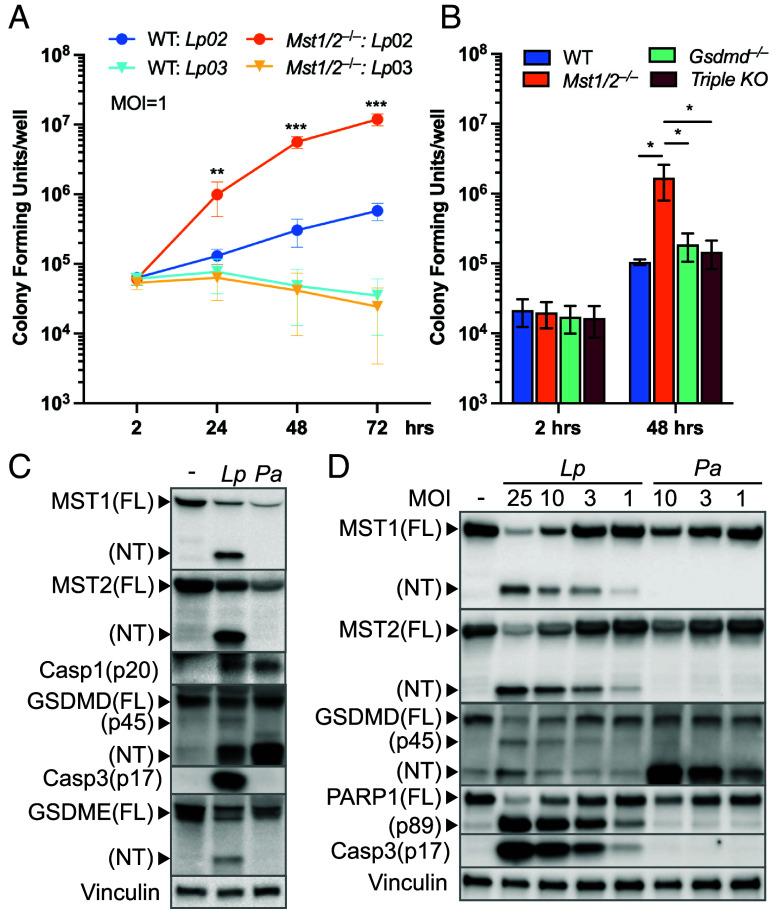
The mammalian Hippo kinases, MST1 and MST2, regulate organ development and suppress tumor formation by balancing cell proliferation and death. In macrophages, inflammasomes detect molecular patterns from invading pathogens or damaged host cells and trigger programmed cell death. In addition to lytic pyroptosis, the signatures associated with apoptosis are induced by inflammasome activation, but how the inflammasomes coordinate different cell death processes remains unclear. Here, we identify the crucial role of MST1/2 in inflammasome-triggered cell death. Macrophages proteolytically convert full-length MST1/2 into the MST1/2 N-terminal fragments (MST1/2-NT) when the NLRC4 inflammasome detects flagellin from the pathogenic bacterium, Legionella pneumophila. Activation of the NLRP3 inflammasome by the damage-associated molecular pattern, extracellular ATP, also produces MST1/2-NT. Caspase-1, the protease activated by these inflammasomes, directly cleaves MST1/2, and blockage of caspase-1 inhibits MST1/2-NT production in macrophages challenged with L. pneumophila. Importantly, MST1/2-NT production is critical for macrophages to trigger a set of death processes associated with apoptosis upon inflammasome activation and knocking out Mst1/2 causes dysregulated gasdermin protein cleavage for pyroptotic death. Furthermore, macrophages lacking MST1/2 have increased susceptibility to virulent L. pneumophila, revealing that the Hippo kinases are important restriction factors against the pathogen. These findings demonstrate that proteolytic cleavage of MST1/2 induced by inflammatory stimuli is an immune pathway to regulate programmed cell death in macrophages and uncover a unique link between the tumor-suppressive Hippo kinases and the inflammasomes in innate immunity.
Keywords: GSDME; STK3; STK4; pattern recognition receptors; type IV secretion.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
