

Life history strategies complement niche partitioning to support the coexistence of closely related Gilliamella species in the bee gut
- PMID: 39893622
- PMCID: PMC11822680
- DOI: 10.1093/ismejo/wraf016
Life history strategies complement niche partitioning to support the coexistence of closely related Gilliamella species in the bee gut
Abstract
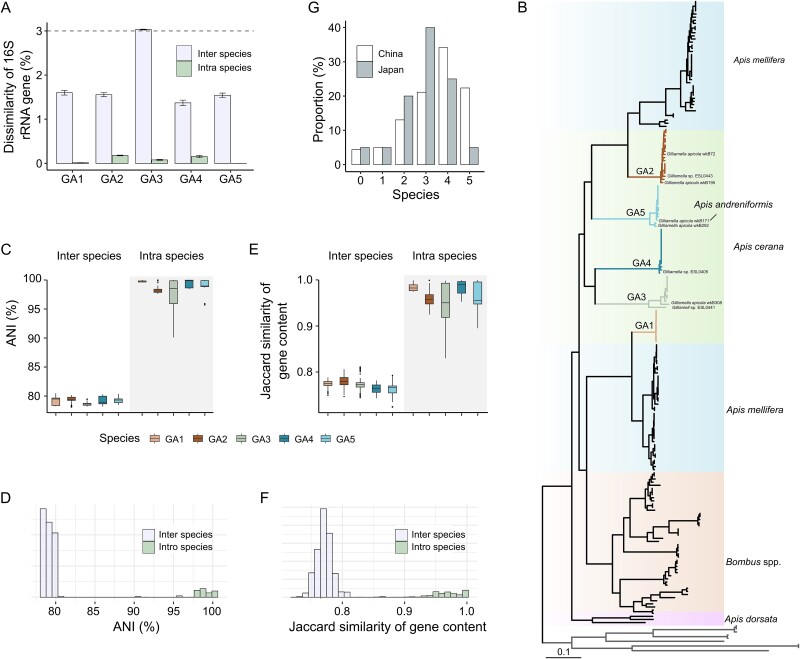
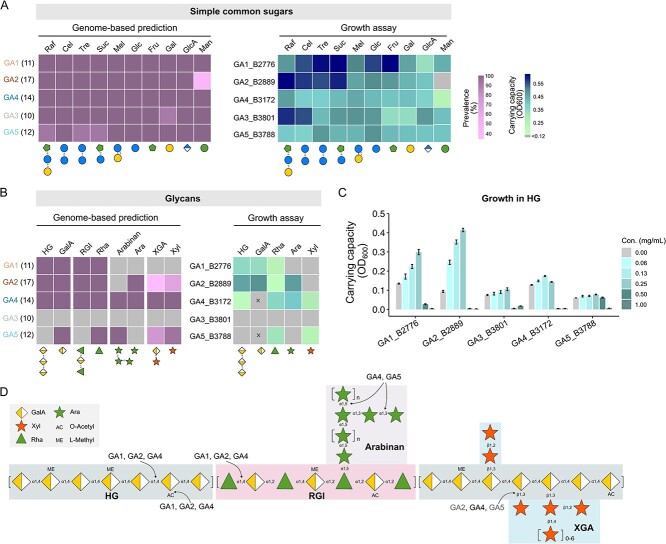
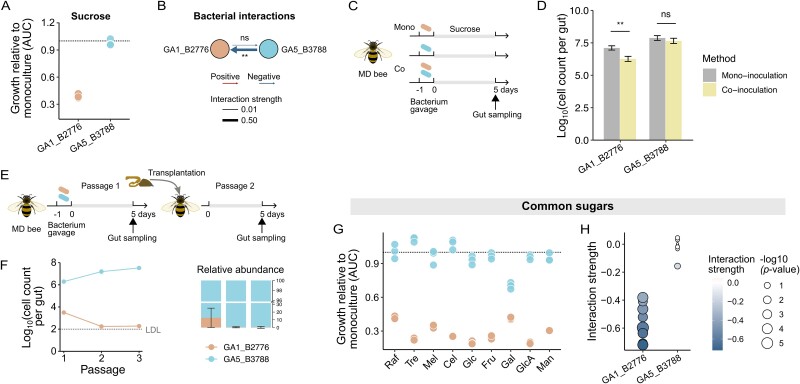
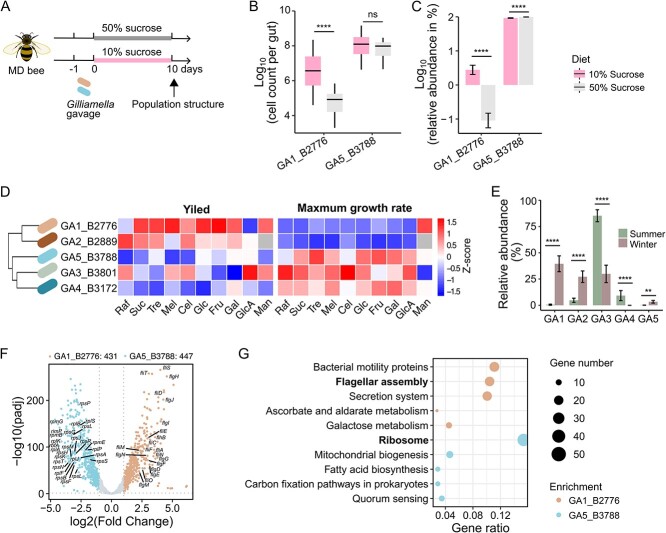
The maintenance of bacterial diversity at both species and strain levels is crucial for the sustainability of honey bee gut microbiota and host health. Periodic or random fluctuation in diet typically alters the metabolic niches available to gut microbes, thereby continuously reshaping bacterial diversity and interspecific interactions. It remains unclear how closely related bacteria adapt to these fluctuations and maintain coexistence within the bee gut. Here, we demonstrate that the five predominant Gilliamella species associated with Apis cerana, a widely distributed Asiatic honey bee, have diverged in carbohydrate metabolism to adapt to distinct nutrient niches driven by dietary fluctuation. Specifically, the glycan-specialists gain improved growth on a pollen-rich diet, but are overall inferior in competition to non-glycan-specialist on either a simple sugar or sugar-pollen diet, when co-inoculated in the bee host and transmitted across generations. Strikingly, despite of their disadvantage in a high-sugar condition, the glycan-specialists are found prevalent in natural A. cerana guts. We further reveal that these bacteria have adopted a life history strategy characterized by high biomass yield on a low-concentration sugar diet, allowing them to thrive under poor nutritional conditions, such as when the bee hosts undergo periodical starvation. Transcriptome analyses indicate that the divergence in life history strategies is attributed to gene expression programming rather than genetic variation. This study highlights the importance of integrative metabolic strategies in carbohydrate utilization, which facilitate the coexistence of closely related Gilliamella species in a changing bee gut environment.
Keywords: r/k selection; gut environmental dynamic; honeybee; interspecific competition; species coexistence.
© The Author(s) 2025. Published by Oxford University Press on behalf of the International Society for Microbial Ecology.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
