

A conserved class of viral RNA structures regulates translation reinitiation through dynamic ribosome interactions
- PMID: 39893634
- PMCID: PMC11921876
- DOI: 10.1016/j.celrep.2025.115236
A conserved class of viral RNA structures regulates translation reinitiation through dynamic ribosome interactions
Abstract
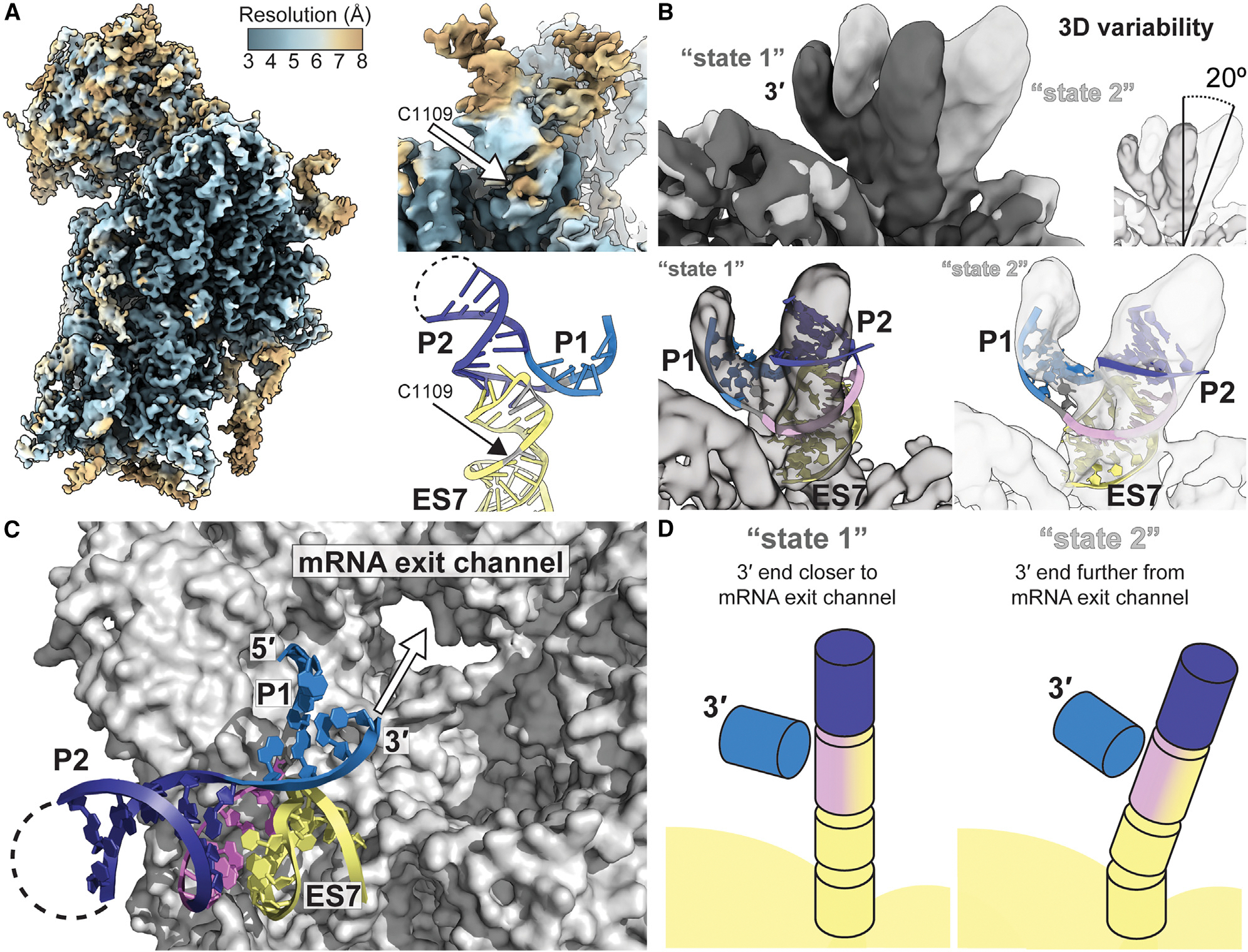
Certain viral RNAs encode proteins downstream of their main open reading frame, expressed through "termination-reinitiation" events. In some cases, structures located upstream of the first stop codon within these viral RNAs bind the ribosome, inhibiting ribosome recycling and inducing reinitiation. We used bioinformatics methods to identify new examples of viral reinitiation-stimulating RNAs and experimentally verified their secondary structure and function. We determined the structure of a representative viral RNA-ribosome complex using cryoelectron microscopy (cryo-EM). 3D classification and variability analyses reveal that the viral RNA structure can sample a range of conformations while remaining tethered to the ribosome, enabling the ribosome to find a reinitiation start site within a limited range of mRNA sequence. Evaluating the conserved features and constraints of this entire RNA class within the context of the cryo-EM reconstruction provides insight into mechanisms enabling reinitiation, a translation regulation strategy employed by many other viral and eukaryotic systems.
Keywords: CP: Molecular biology; RNA structure; cryo-EM; reinitiation; ribosome; viral RNA.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Update of
-
A conserved class of viral RNA structures regulate translation reinitiation through dynamic ribosome interactions.bioRxiv [Preprint]. 2023 Sep 29:2023.09.29.560040. doi: 10.1101/2023.09.29.560040. bioRxiv. 2023. Update in: Cell Rep. 2025 Feb 25;44(2):115236. doi: 10.1016/j.celrep.2025.115236. PMID: 37808774 Free PMC article. Updated. Preprint.
References
-
- Aitken CE, and Lorsch JR (2012). A mechanistic overview of translation initiation in eukaryotes. Nat. Struct. Mol. Biol. 19, 568–576. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
