

This is a preprint.
Markovian State Models uncover Casein Kinase 1 dynamics that govern circadian period
- PMID: 39896482
- PMCID: PMC11785140
- DOI: 10.1101/2025.01.17.633651
Markovian State Models uncover Casein Kinase 1 dynamics that govern circadian period
Abstract
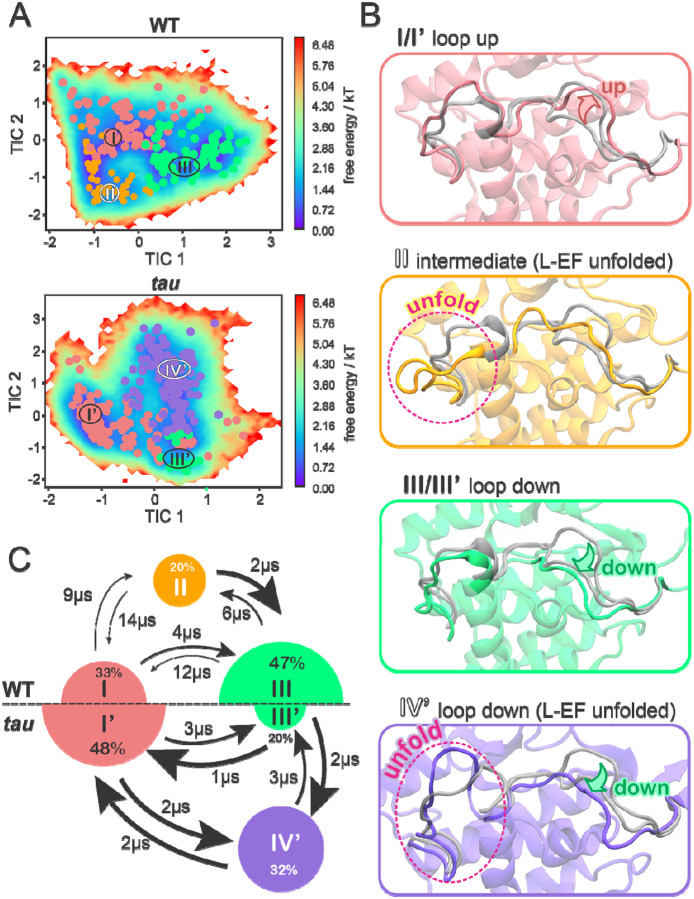
Circadian rhythms in mammals are tightly regulated through phosphorylation of Period (PER) proteins by Casein Kinase 1 (CK1, subtypes δ and ε). CK1 acts on at least two different regions of PER with opposing effects: phosphorylation of phosphodegron (pD) regions leads to PER degradation, while phosphorylation of the Familial Advanced Sleep Phase (FASP) region leads to PER stabilization. To investigate how substrate selectivity is encoded by the conformational dynamics of CK1, we performed a large set of independent molecular dynamics (MD) simulations of wildtype CK1 and the tau mutant (R178C) that biases kinase activity toward a pD. We used Markovian State Models (MSMs) to integrate the simulations into a single model of the conformational landscape of CK1 and used Gaussian accelerated molecular dynamics (GaMD) to build the first molecular model of CK1 and the unphosphorylated FASP motif. Together, these findings provide a mechanistic view of CK1, establishing how the activation loop acts as a key molecular switch to control substrate selectivity. We show that the tau mutant favors an alternative conformation of the activation loop and significantly accelerates the dynamics of CK1. This reshapes the binding cleft in a way that impairs FASP binding and would ultimately lead to PER destabilization and shorter circadian periods. Finally, we identified an allosteric pocket that could be targeted to bias this molecular switch. Our integrated approach offers a detailed model of CK1's conformational landscape and its relevance to normal, mutant, and druggable circadian timekeeping.
Conflict of interest statement
Declaration of interests The authors do not declare any conflicts of interest.
Figures






References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials