

This is a preprint.
ADAR1 haploinsufficiency and sustained picornaviral RdRp dsRNA synthesis synergize to dysregulate RNA editing and cause multi-system interferonopathy
- PMID: 39896491
- PMCID: PMC11785089
- DOI: 10.1101/2025.01.21.634124
ADAR1 haploinsufficiency and sustained picornaviral RdRp dsRNA synthesis synergize to dysregulate RNA editing and cause multi-system interferonopathy
Update in
-
ADAR1 haploinsufficiency and sustained picornaviral RdRp dsRNA synthesis synergize to dysregulate RNA editing and cause multi-system interferonopathy.mBio. 2025 Aug 13;16(8):e0149225. doi: 10.1128/mbio.01492-25. Epub 2025 Jul 22. mBio. 2025. PMID: 40693792 Free PMC article.
Abstract
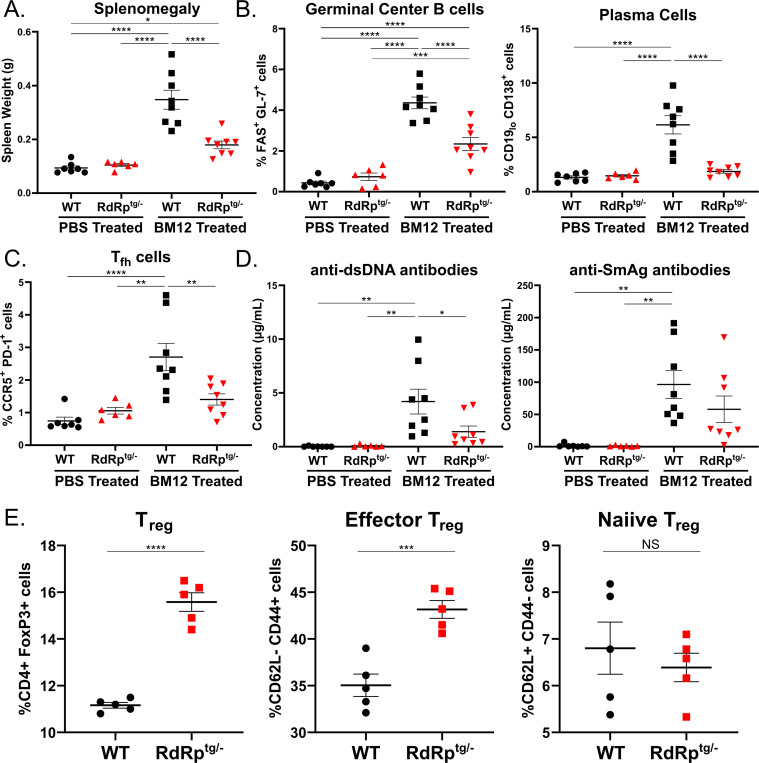
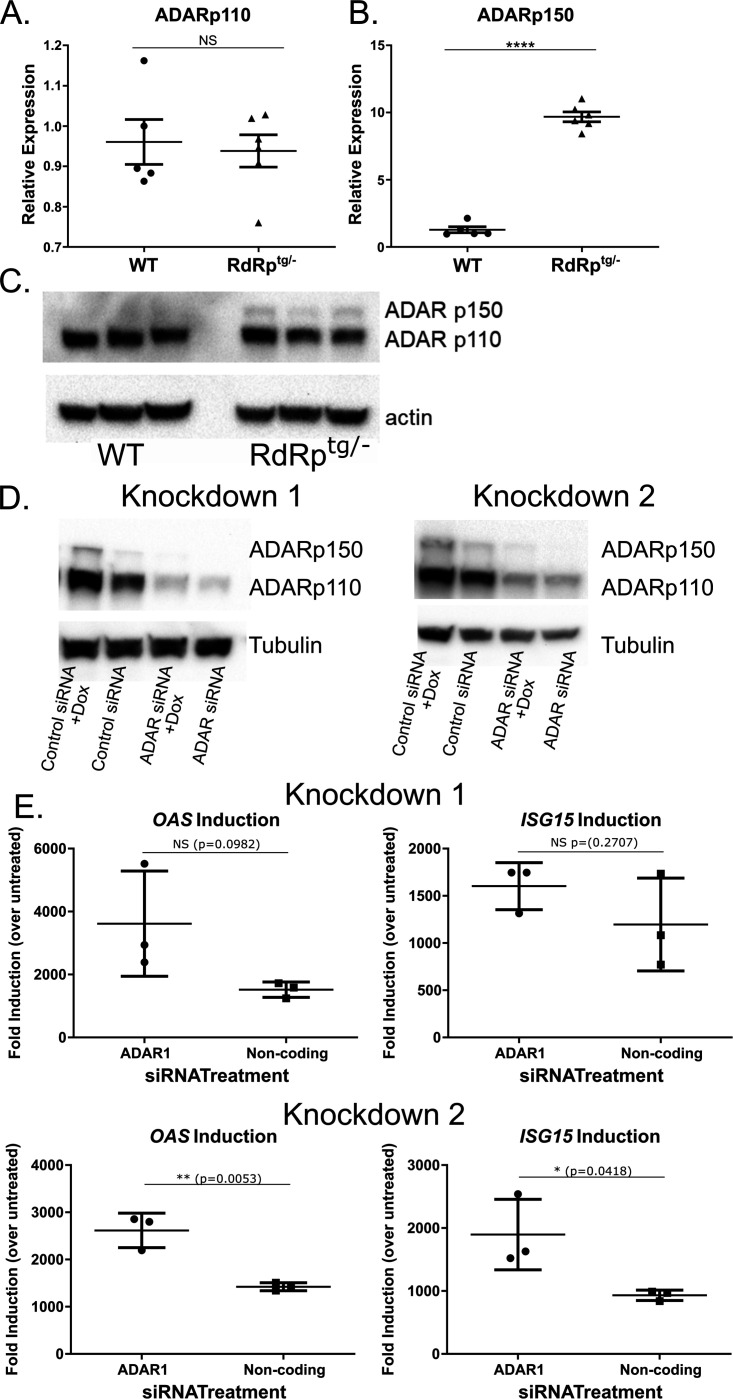
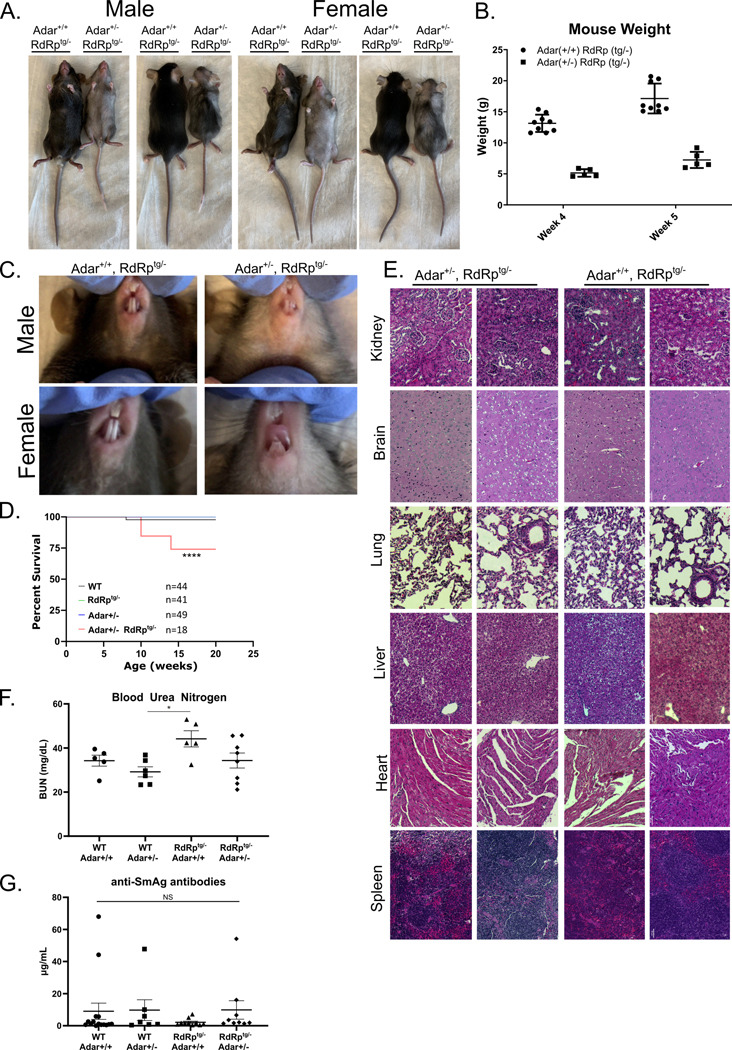
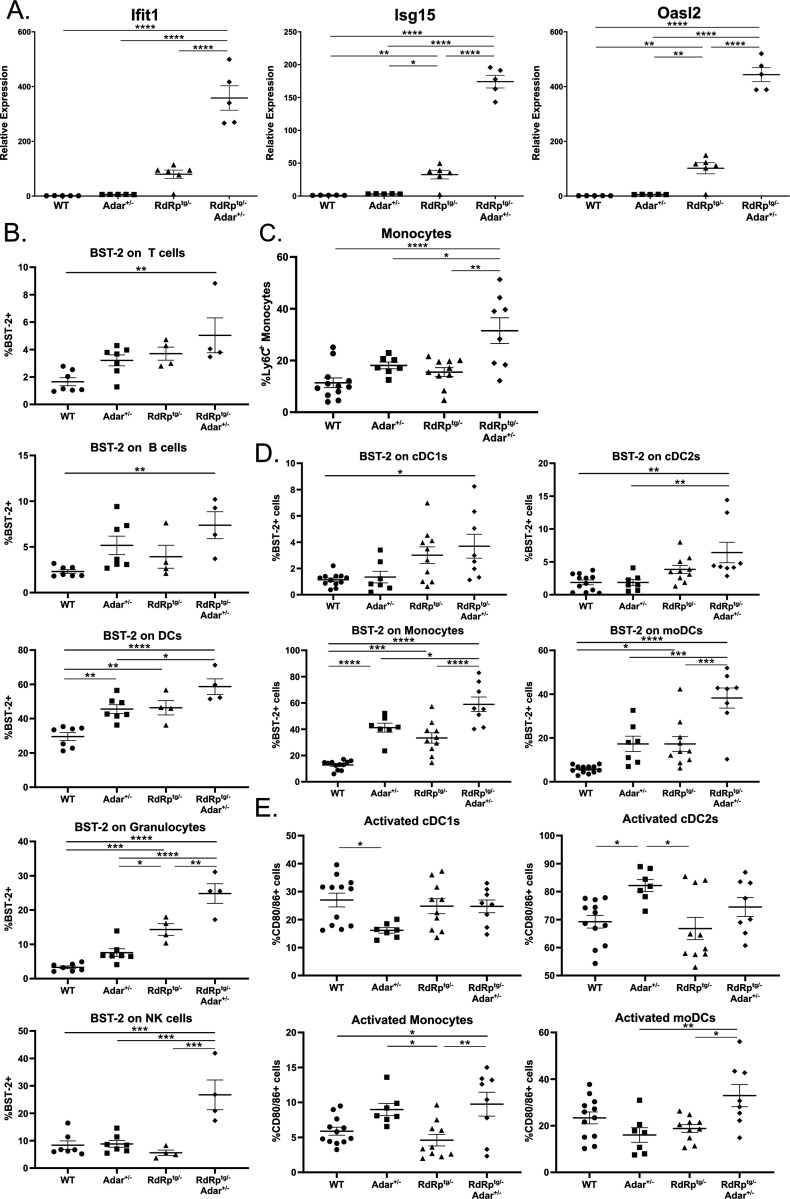
Sensing of viral double-stranded RNA by MDA5 triggers abundant but transient interferon-stimulated gene (ISGs) expression. If dsRNA synthesis is made persistent by transgenically expressing a picornaviral RNA-dependent RNA polymerase (RdRp) in mice, lifelong MDA5-MAVS pathway activation and marked, global ISG upregulation result. This confers robust protection from viral diseases but in contrast to numerous other chronic MDA5 hyperactivation states, the mice suffer no autoimmune or other health consequences. Here we find they further confound expectations by being resistant to a strong autoimmunity (lupus) provocation. However, knockout of one allele of Adar breaks the autoinflammation-protected state of RdRptg mice and results in a severe disease that resembles interferonopathies caused by MDA5 gain-of-function protein mutations. Adar +/- mice are healthy but Adar +/- RdRptg mice have shortened lifespan, stunted growth, premature fur graying, poorly developed teeth, skeletal abnormalities, and extreme ISG elevations. A-to-I edits are both abnormally distributed and increased (numbers of genes and sites). These results, with a nucleic acid-triggered and MDA5-wild type model, illuminate the ADAR1-MDA5 axis in the regulation of innate immunity and establish that viral polymerase-sourced dsRNA can drive autoinflammatory disease pathogenesis.
Keywords: ADAR1; MDA5; RdRp; autoimmunity; autoinflammation; innate antiviral immunity; interferonopathy; lupus; picornavirus.
Figures



References
-
- Hornung V, Ellegast J, Kim S, Brzozka K, Jung A, Kato H, Poeck H, Akira S, Conzelmann KK, Schlee M, Endres S, Hartmann G. 2006. 5’-Triphosphate RNA is the ligand for RIG-I. Science 314:994–7. - PubMed
-
- Kato H, Takeuchi O, Mikamo-Satoh E, Hirai R, Kawai T, Matsushita K, Hiiragi A, Dermody TS, Fujita T, Akira S. 2008. Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J Exp Med 205:1601–10. - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous