

This is a preprint.
Mettl15-Mettl17 modulates the transition from early to late pre-mitoribosome
- PMID: 39896671
- PMCID: PMC11785013
- DOI: 10.1101/2024.12.18.629302
Mettl15-Mettl17 modulates the transition from early to late pre-mitoribosome
Update in
-
Mettl15-Mettl17 modulates the transition from early to late pre-mitoribosome.Structure. 2025 Nov 6;33(11):1904-1915.e3. doi: 10.1016/j.str.2025.08.002. Epub 2025 Aug 28. Structure. 2025. PMID: 40882632
Abstract
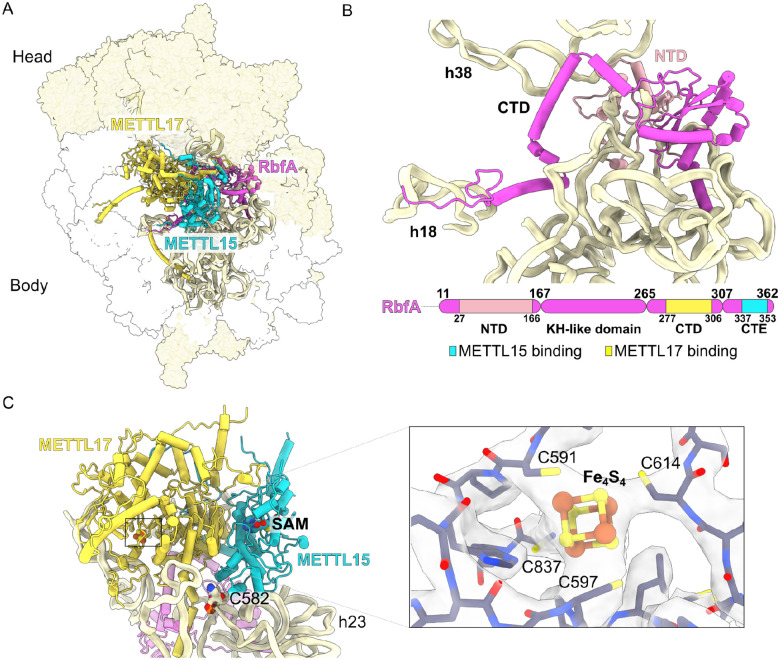
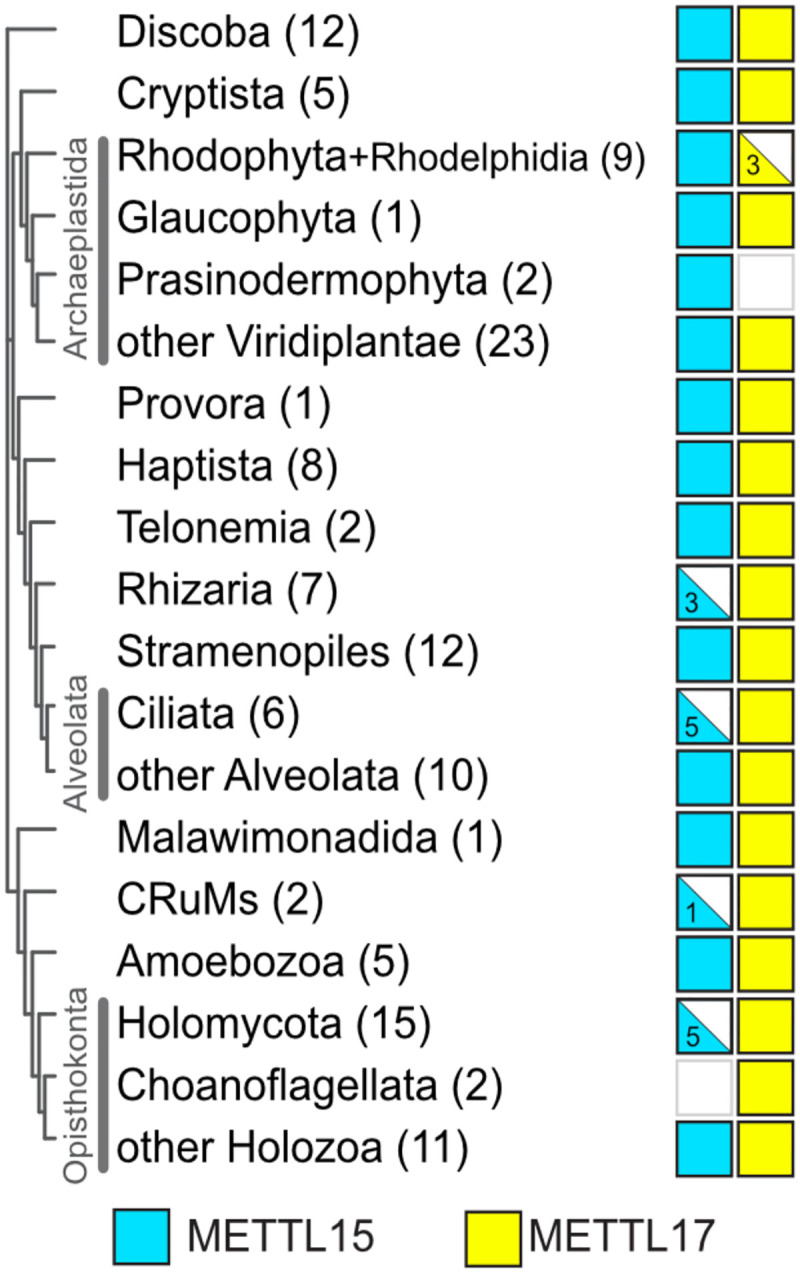
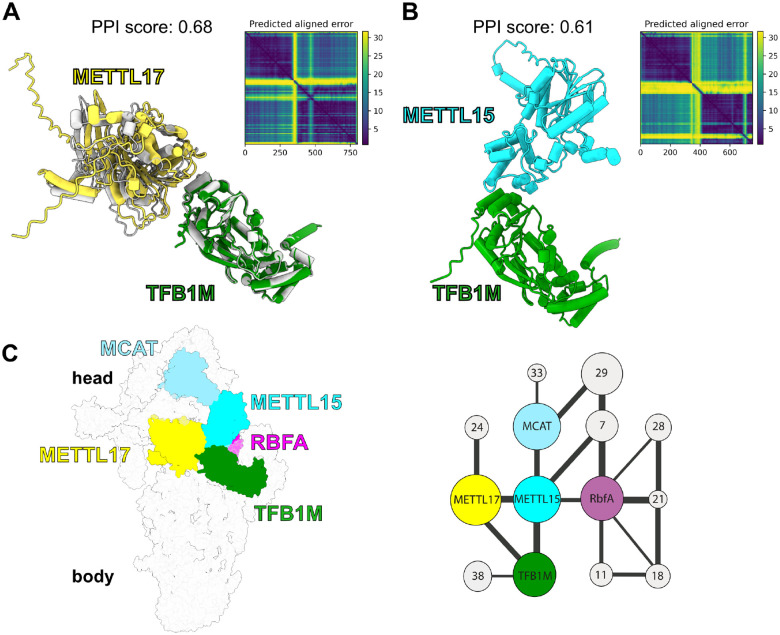
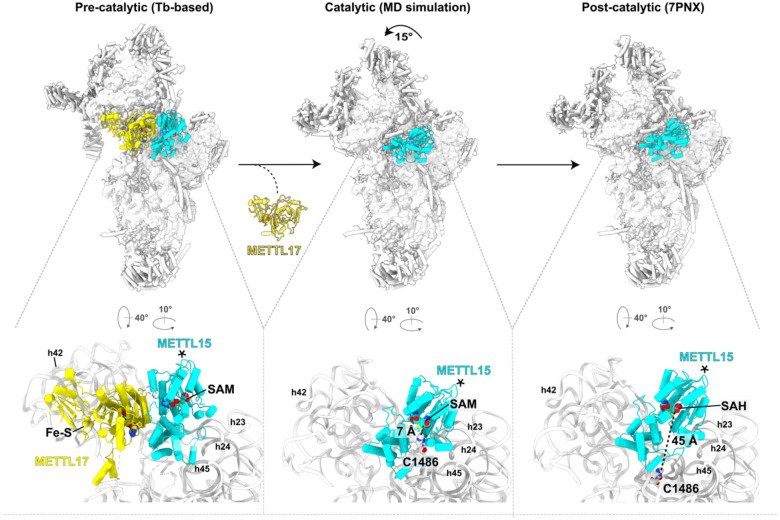
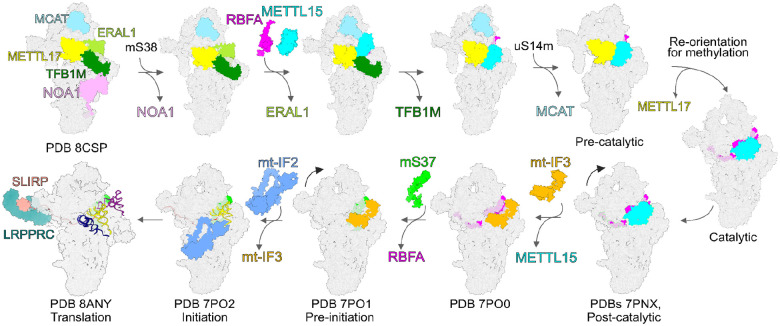
The assembly of the mitoribosomal small subunit involves folding and modification of rRNA, and its association with mitoribosomal proteins. This process is assisted by a dynamic network of assembly factors. Conserved methyltransferases Mettl15 and Mettl17 act on the solvent-exposed surface of rRNA. Binding of Mettl17 is associated with the early assembly stage, whereas Mettl15 is involved in the late stage, but the mechanism of transition between the two was unclear. Here, we integrate structural data from Trypanosoma brucei with mammalian homologs and molecular dynamics simulations. We reveal how the interplay of Mettl15 and Mettl17 in intermediate steps links the distinct stages of small subunit assembly. The analysis suggests a model wherein Mettl17 acts as a platform for Mettl15 recruitment. Subsequent release of Mettl17 allows a conformational change of Mettl15 for substrate recognition. Upon methylation, Mettl15 adopts a loosely bound state which ultimately leads to its replacement by initiation factors, concluding the assembly. Together, our results indicate that assembly factors Mettl15 and Mettl17 cooperate to regulate the biogenesis process, and present a structural data resource for understanding molecular adaptations of assembly factors in mitoribosome.
Figures

References
-
- Ott M., Amunts A. & Brown A. Organization and Regulation of Mitochondrial Protein Synthesis. Annu Rev Biochem 85, 77–101 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources