

Structural insights into dynamics of the BMV TLS aminoacylation
- PMID: 39900568
- PMCID: PMC11791061
- DOI: 10.1038/s41467-025-56612-4
Structural insights into dynamics of the BMV TLS aminoacylation
Abstract
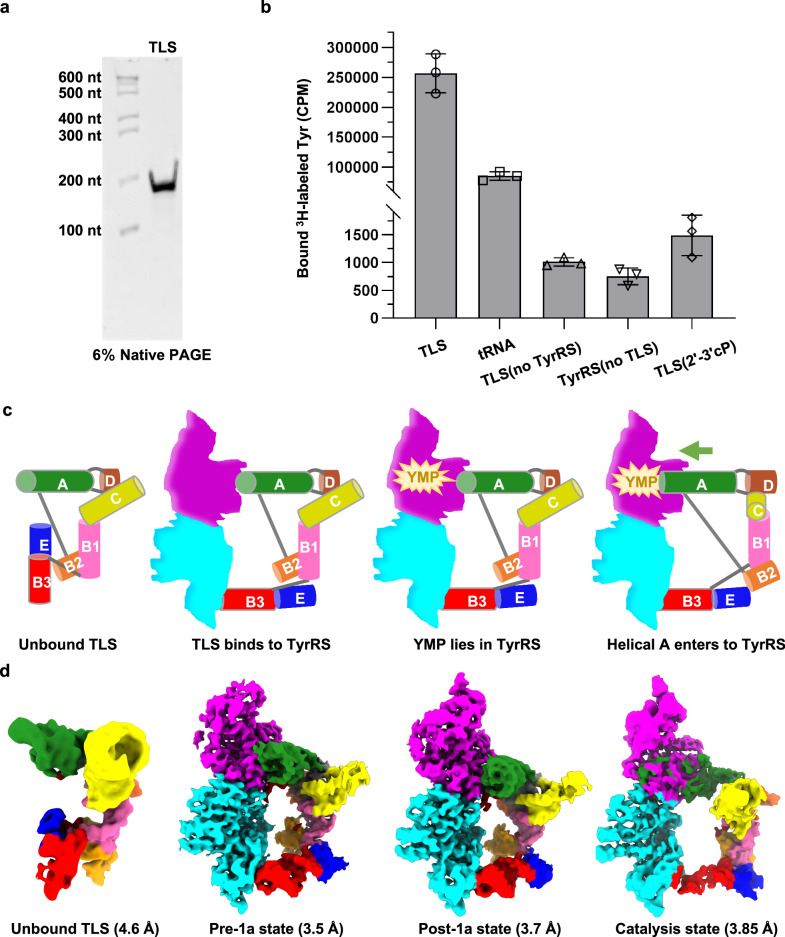
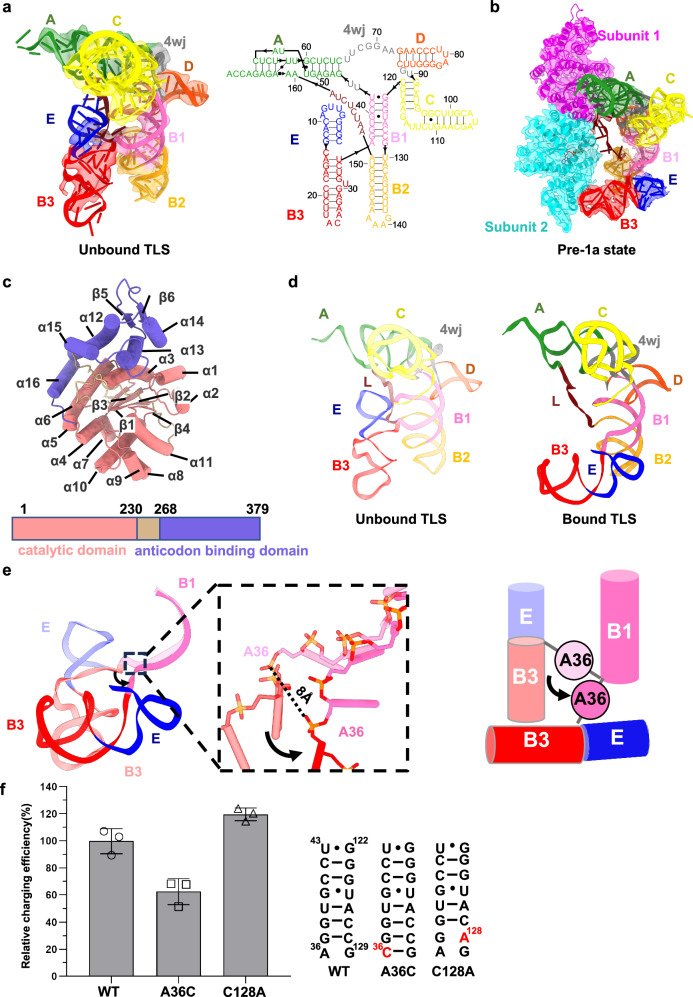
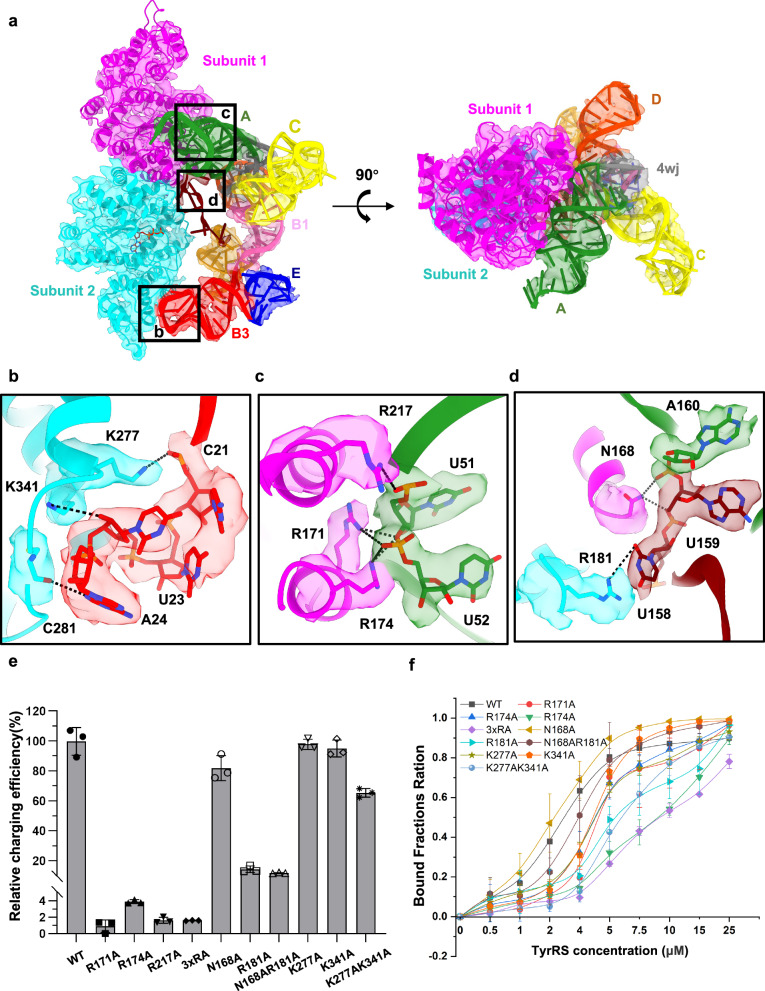
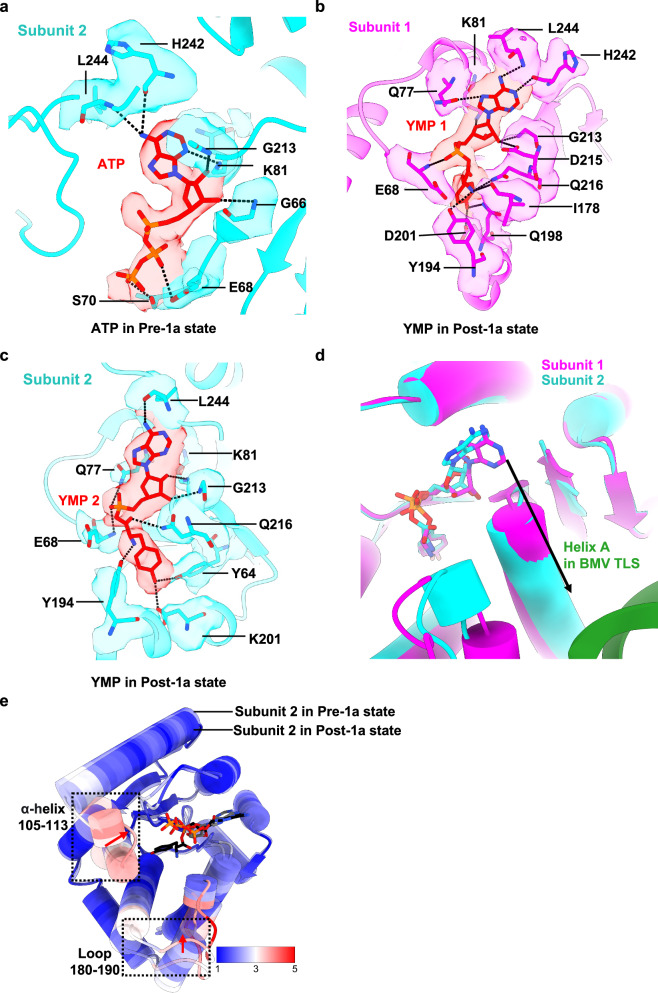
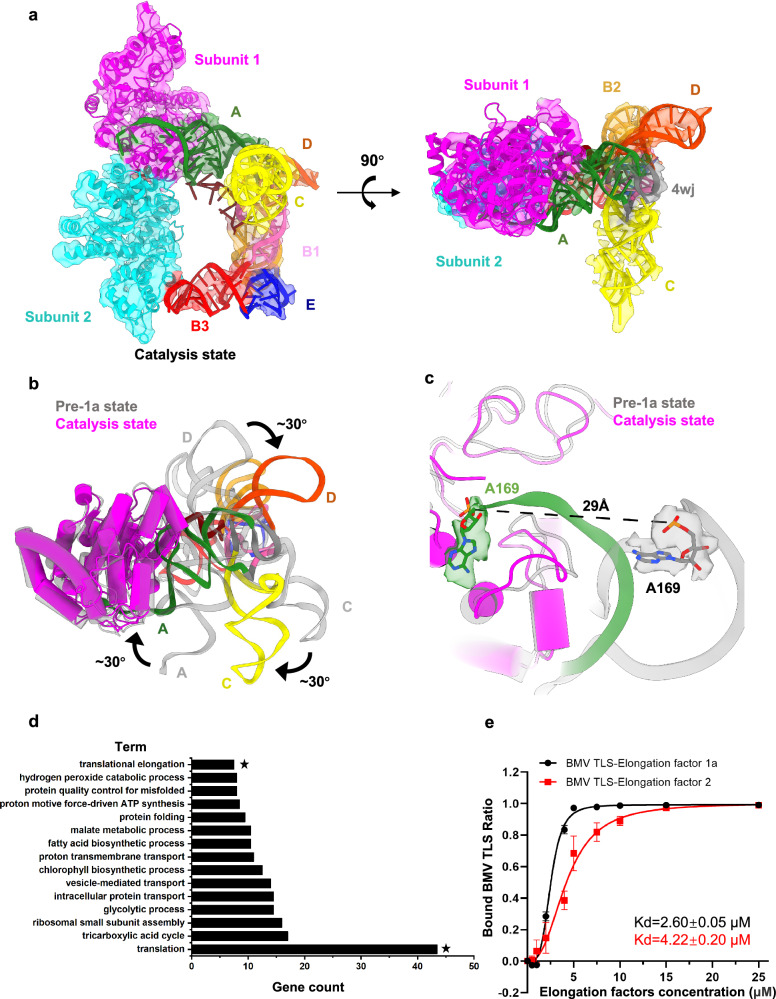
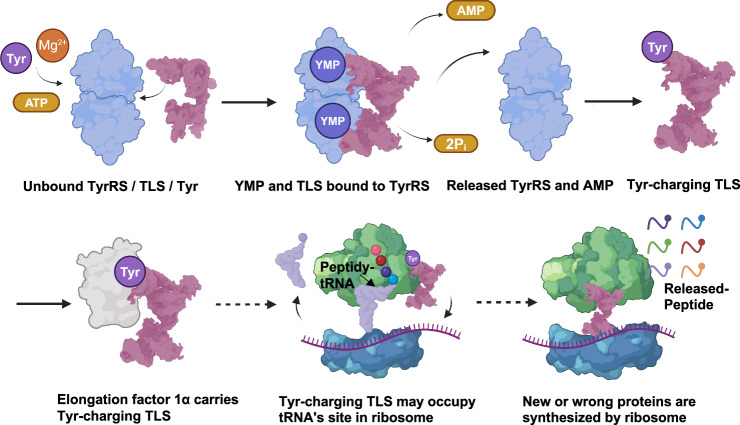
Brome Mosaic Virus (BMV) utilizes a tRNA-like structure (TLS) within its 3' untranslated region to mimic host tRNA functions, aiding aminoacylation and viral replication. This study explores the structural dynamics of BMV TLS interacting with tyrosyl-tRNA synthetase (TyrRS) during aminoacylation. Using cryo-EM, we capture multiple states of the TLS-TyrRS complex, including unbound TLS, pre-1a, post-1a, and catalysis states, with resolutions of 4.6 Å, 3.5 Å, 3.7 Å, and 3.85 Å, respectively. These structural comparisons indicate dynamic changes in both TLS and TyrRS. Upon binding, TLS undergoes dynamic rearrangements, particularly with helices B3 and E pivoting, mediated by the unpaired A36 residue, ensuring effective recognition by TyrRS. The dynamic changes also include a more compact arrangement in the catalytic center of TyrRS and the insertion of 3' CCA end into the enzyme's active site, facilitating two-steps aminoacylation. Enzymatic assays further demonstrated the functional importance of TLS-TyrRS interactions, with mutations in key residues significantly impacting aminoacylation efficiency. Furthermore, Electrophoretic Mobility Shift Assay (EMSA) demonstrated that BMV TLS binds elongation factors EF1α and EF2, suggesting a multifaceted strategy to exploit host translational machinery. These findings not only enhance our knowledge of virus-host interactions but also offer potential targets for antiviral drug development.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
