

Super-enhancers and efficacy of triptolide in small cell carcinoma of the ovary hypercalcemic type
- PMID: 39906560
- PMCID: PMC11791298
- DOI: 10.1016/j.isci.2025.111770
Super-enhancers and efficacy of triptolide in small cell carcinoma of the ovary hypercalcemic type
Abstract
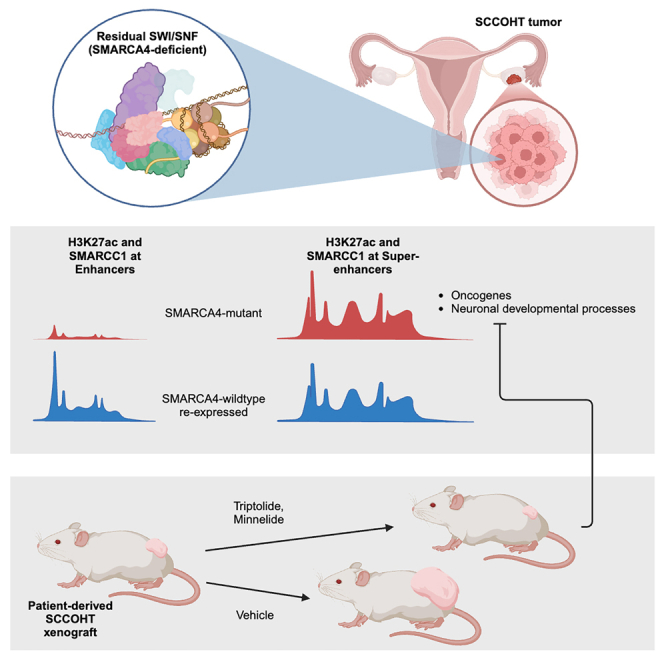
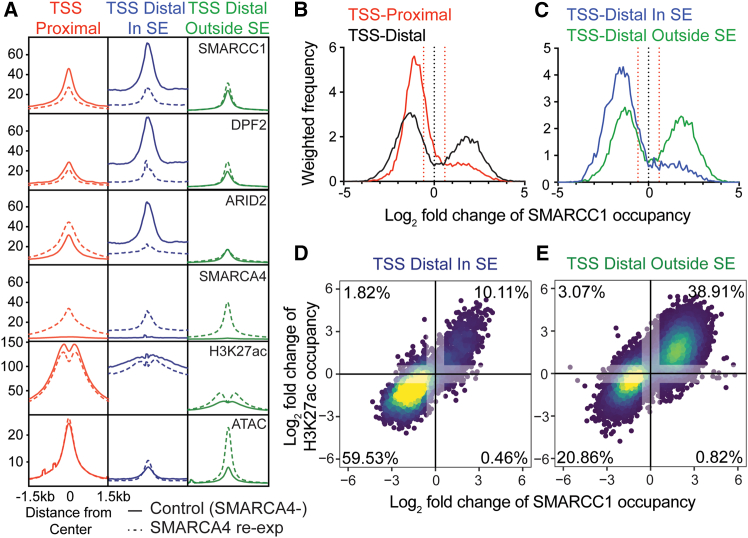
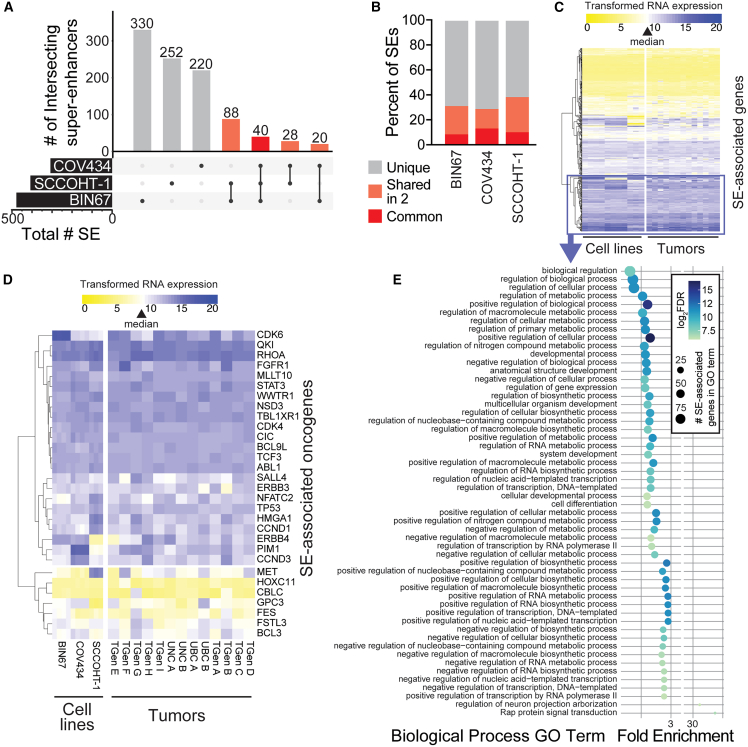
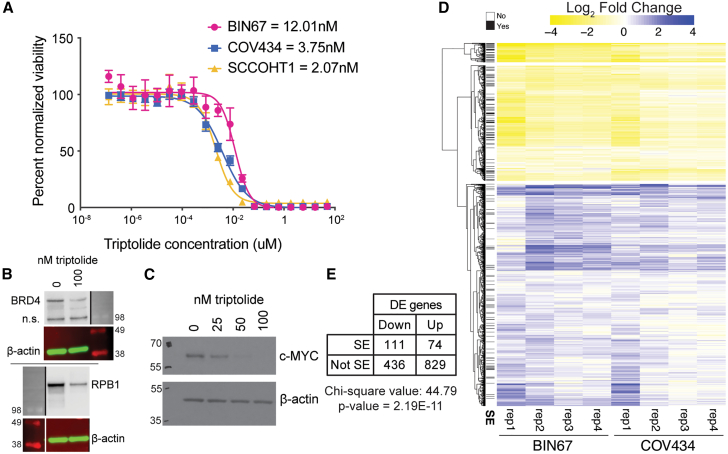
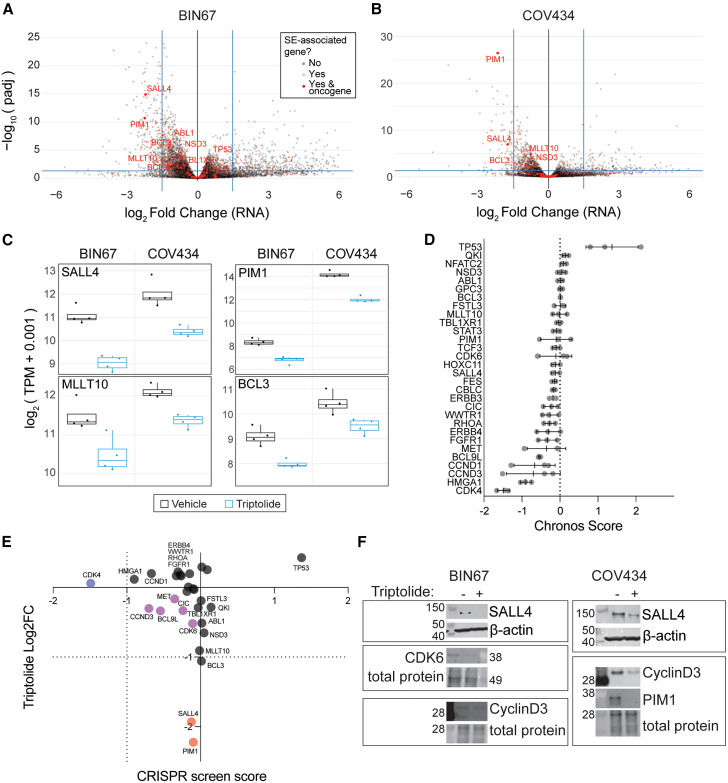
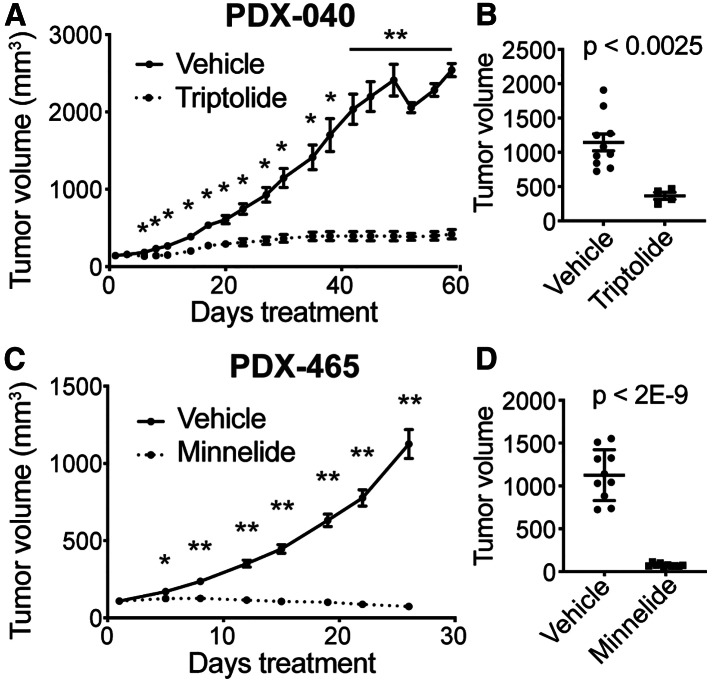
Small cell carcinoma of the ovary-hypercalcemic type (SCCOHT) is a rare ovarian cancer affecting young females and is driven by the loss of both SWI/SNF ATPases SMARCA4 and SMARCA2. As loss of SWI/SNF alters enhancers, we hypothesized that super-enhancers, which regulate oncogene expression in cancer, are disparately impacted by SWI/SNF loss. We discovered differences between SWI/SNF occupancy at enhancers vs. super-enhancers. SCCOHT super-enhancer target genes were enriched in developmental processes, most notably nervous system development. This may further support neuronal cell-of-origin previously proposed. We found high sensitivity of SCCOHT cell lines to triptolide. Triptolide inhibits expression of many super-enhancer-associated genes, including oncogenes. SALL4 expression is decreased by triptolide and is highly expressed in SCCOHT tumors. In patient-derived xenograft models, triptolide and prodrug minnelide effectively inhibit tumor growth. These results reveal unique features of super-enhancers in SCCOHT, which may be one mechanism through which triptolide has high activity in these tumors.
Keywords: Cancer; Complex system biology; Molecular biology.
© 2025 The Authors.
Conflict of interest statement
Since their contributions to this manuscript, several authors have new institutional affiliations not listed in the author affiliations: William Selleck - current affiliation: CellProtein Sciences, LLC (consultant); Shawn Striker - current affiliation: The Ohio State University; Nicolle Hipschman - current affliation: University of Arizona; Salvatore J Facista - current affiliation: Anivive; William P. D. Hendricks - current affiliation: Actual Labs Advisory (advisor and consultant), Vetted Capital (Venture Partner & Scientific Advisor), MI:RNA Diagnostics (Product Development Advisor); Krystal A. Orlando - current affiliation: National Institute of Environmental Health Sciences, National Institutes of Health; Elizabeth A. Raupach - current affiliation: Mayo Clinic in Arizona.
Figures

References
-
- Witkowski L., Goudie C., Ramos P., Boshari T., Brunet J.-S., Karnezis A.N., Longy M., Knost J.A., Saloustros E., McCluggage W.G., et al. The influence of clinical and genetic factors on patient outcome in small cell carcinoma of the ovary, hypercalcemic type. Gynecol. Oncol. 2016;141:454–460. doi: 10.1016/j.ygyno.2016.03.013. - DOI - PMC - PubMed
-
- Tischkowitz M., Huang S., Banerjee S., Hague J., Hendricks W.P.D., Huntsman D.G., Lang J.D., Orlando K.A., Oza A.M., Pautier P., et al. Small-Cell Carcinoma of the Ovary, Hypercalcemic Type-Genetics, New Treatment Targets, and Current Management Guidelines. Clin. Cancer Res. 2020;26:3908–3917. doi: 10.1158/1078-0432.CCR-19-3797. - DOI - PMC - PubMed
-
- Witkowski L., Carrot-Zhang J., Albrecht S., Fahiminiya S., Hamel N., Tomiak E., Grynspan D., Saloustros E., Nadaf J., Rivera B., et al. Germline and somatic SMARCA4 mutations characterize small cell carcinoma of the ovary, hypercalcemic type. Nat. Genet. 2014;46:438–443. doi: 10.1038/ng.2931. - DOI - PubMed
-
- Karnezis A.N., Wang Y., Ramos P., Hendricks W.P., Oliva E., D’Angelo E., Prat J., Nucci M.R., Nielsen T.O., Chow C., et al. Dual loss of the SWI/SNF complex ATPases SMARCA4/BRG1 and SMARCA2/BRM is highly sensitive and specific for small cell carcinoma of the ovary, hypercalcaemic type. J. Pathol. 2016;238:389–400. doi: 10.1002/path.4633. - DOI - PMC - PubMed
-
- Ramos P., Karnezis A.N., Hendricks W.P.D., Wang Y., Tembe W., Zismann V.L., Legendre C., Liang W.S., Russell M.L., Craig D.W., et al. Loss of the tumor suppressor SMARCA4 in small cell carcinoma of the ovary, hypercalcemic type (SCCOHT) Rare Dis. 2014;2 doi: 10.4161/2167549X.2014.967148. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
