

Multilevel plasticity and altered glycosylation drive aggressiveness in hypoxic and glucose-deprived bladder cancer cells
- PMID: 39906564
- PMCID: PMC11791300
- DOI: 10.1016/j.isci.2025.111758
Multilevel plasticity and altered glycosylation drive aggressiveness in hypoxic and glucose-deprived bladder cancer cells
Abstract
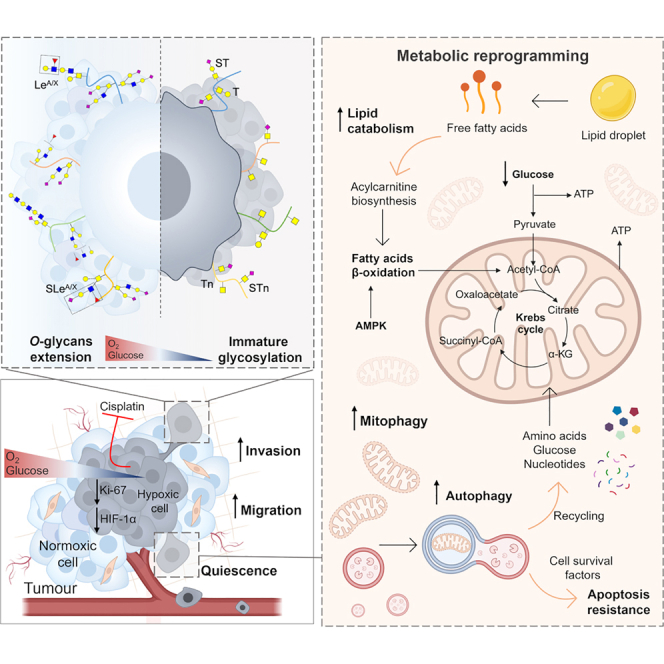
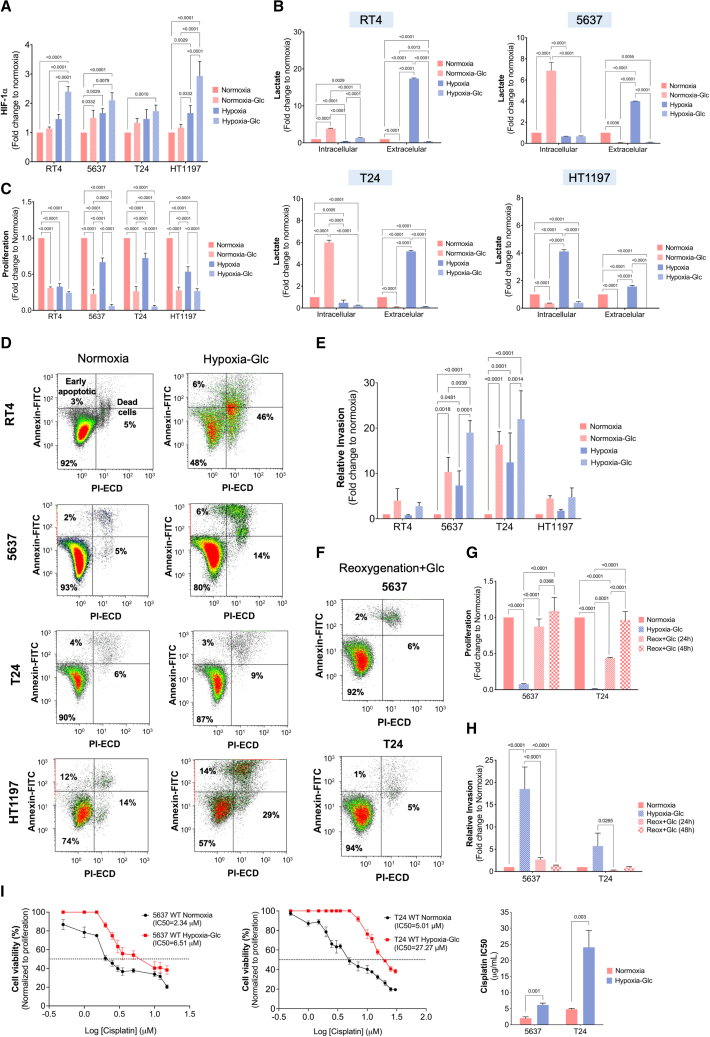
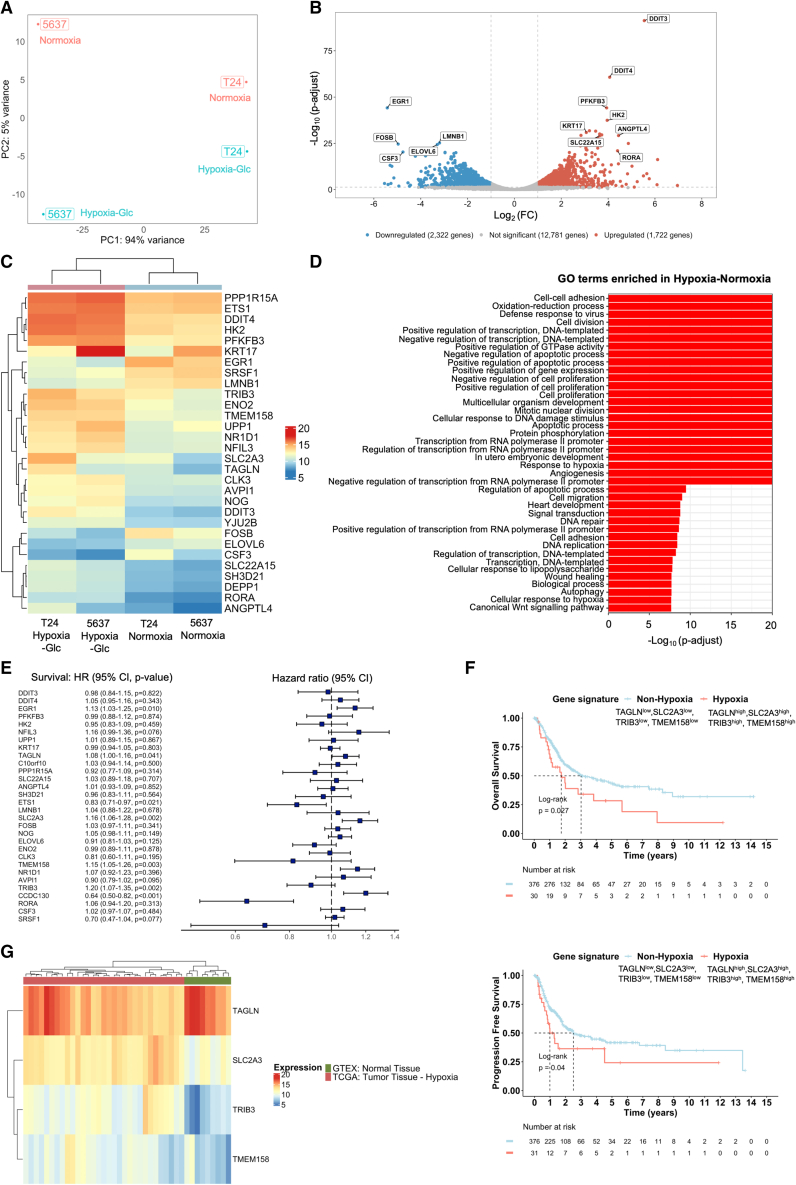
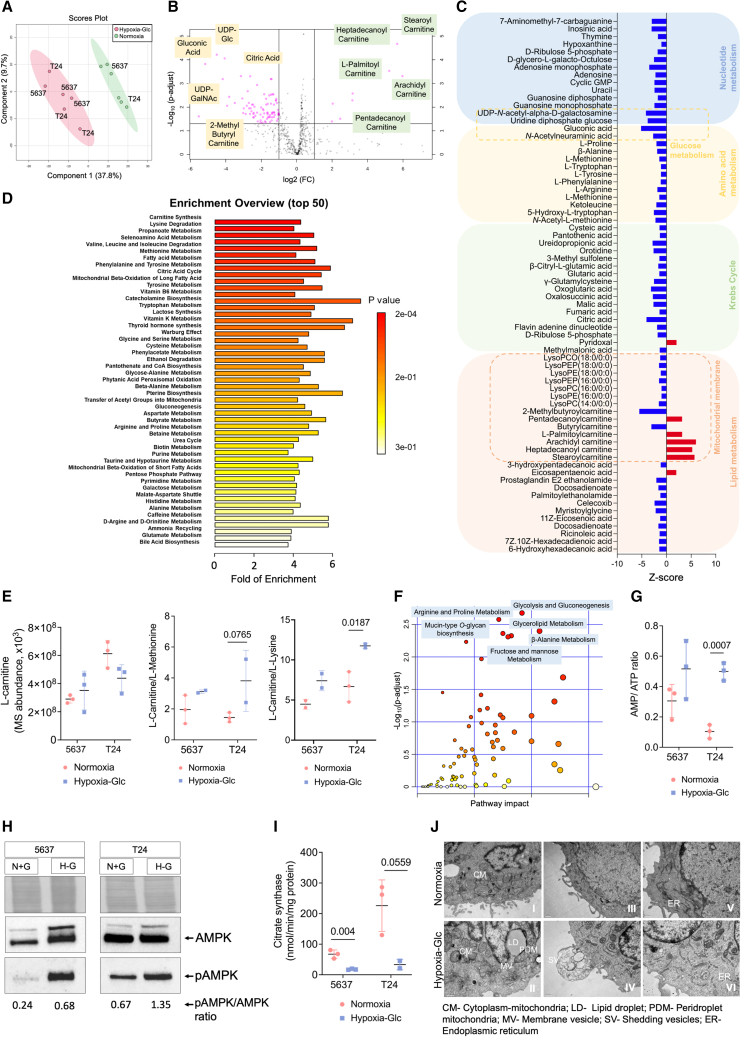
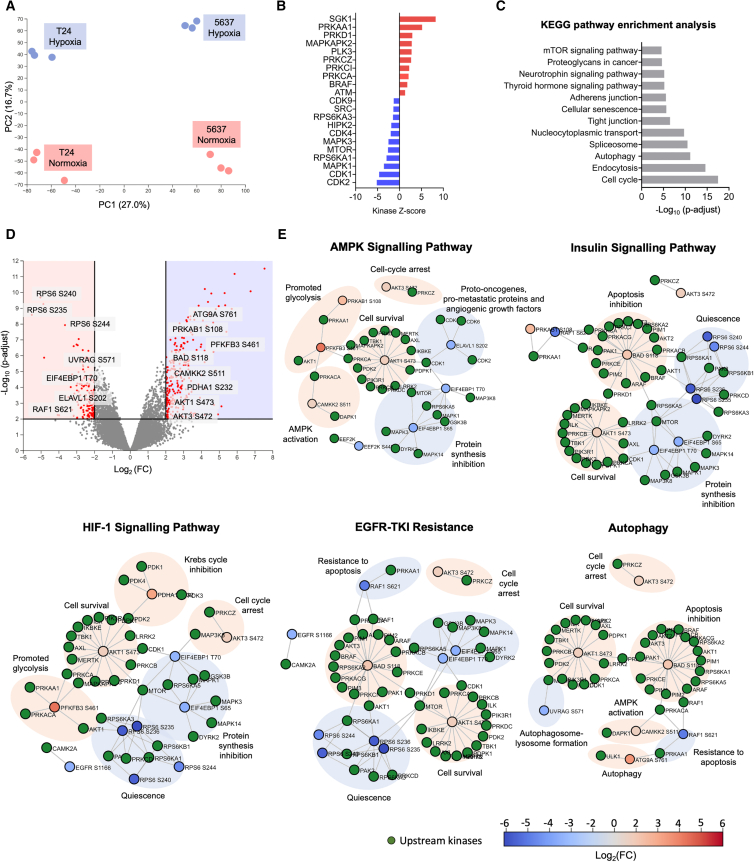
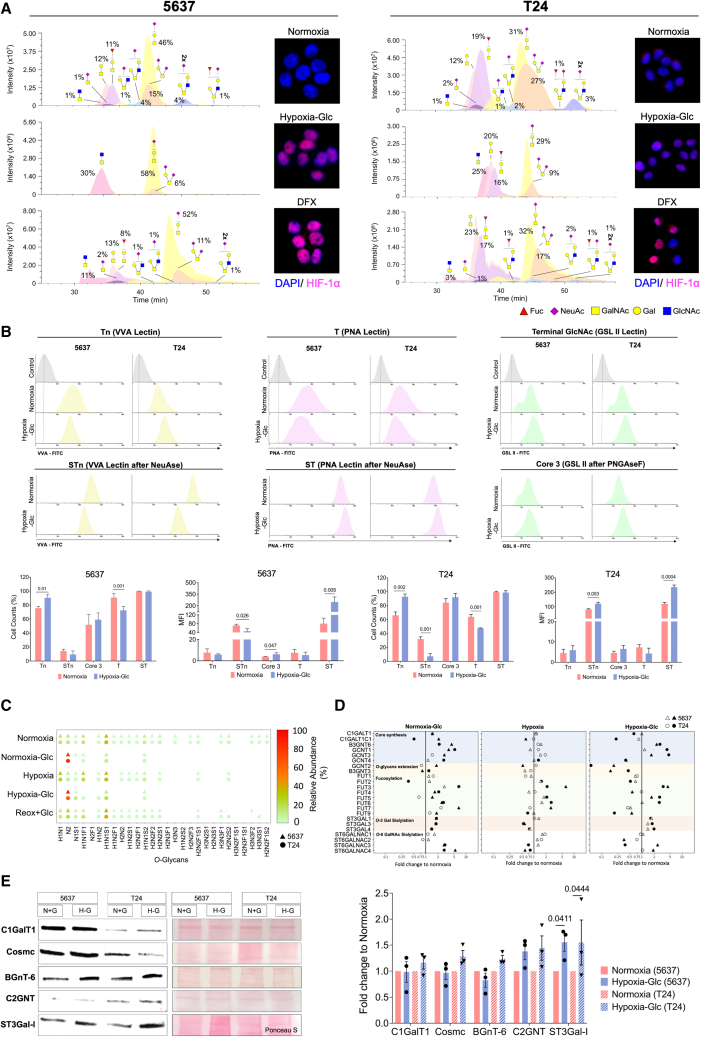
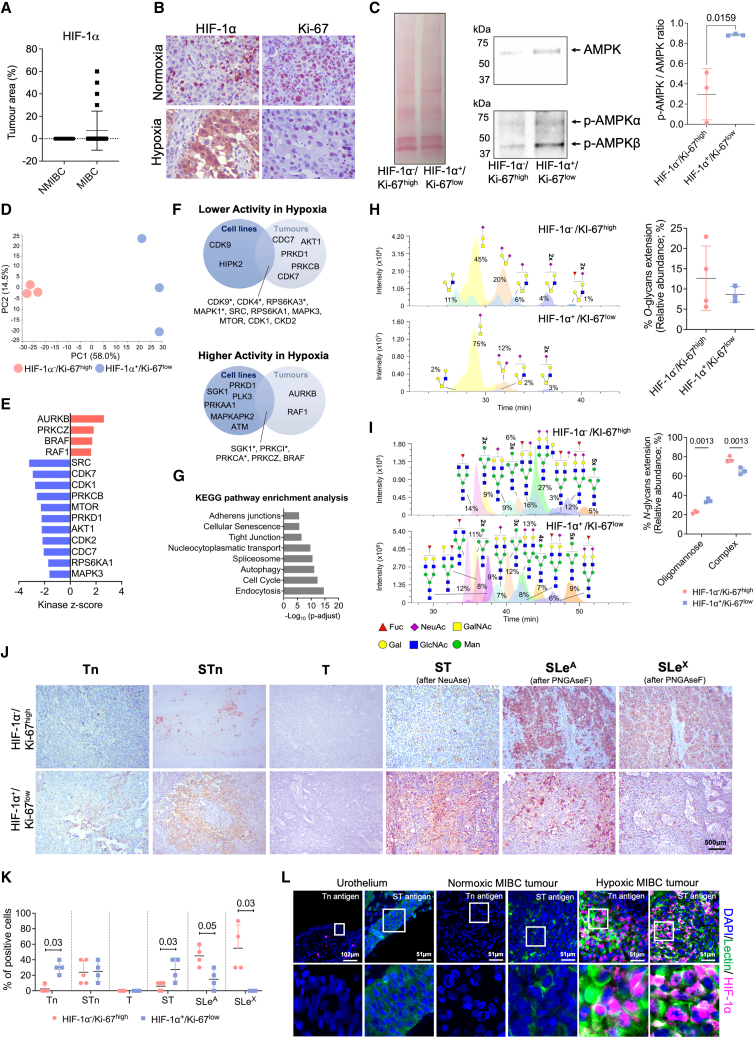
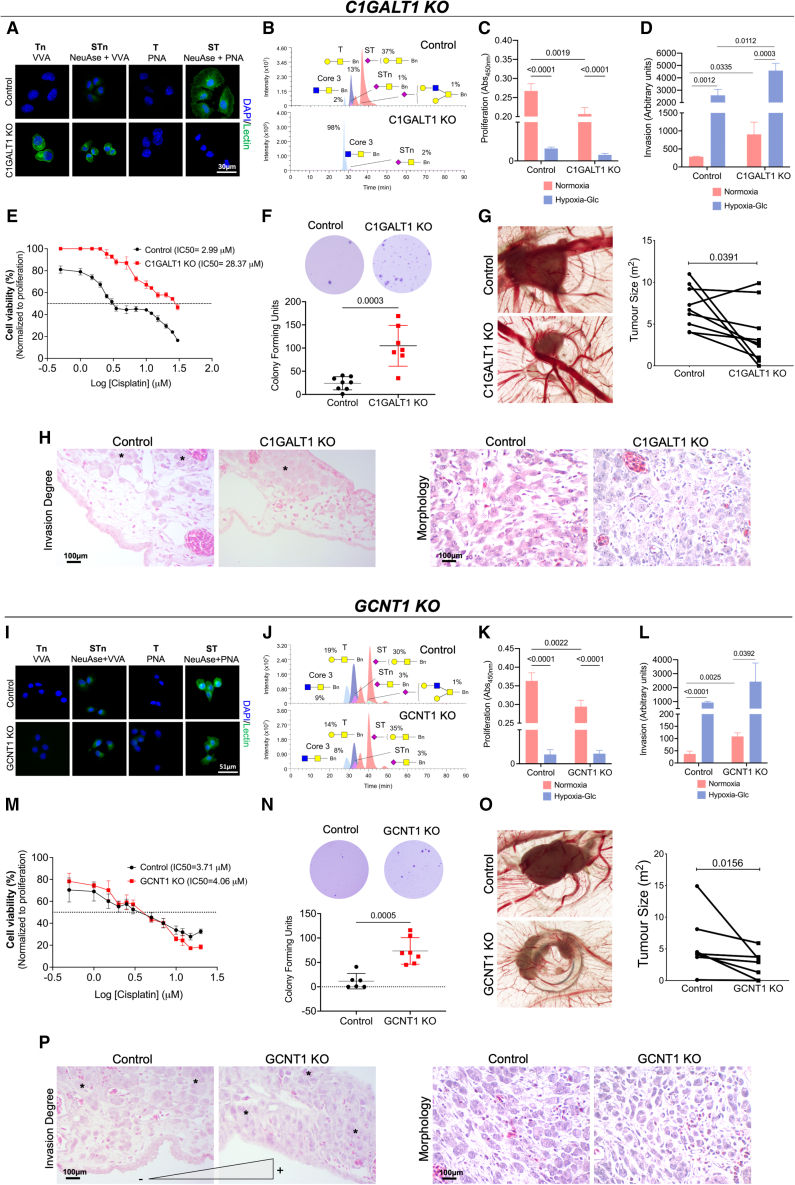
Bladder tumors with aggressive characteristics often present microenvironmental niches marked by low oxygen levels (hypoxia) and limited glucose supply due to inadequate vascularization. The molecular mechanisms facilitating cellular adaptation to these stimuli remain largely elusive. Employing a multi-omics approach, we discovered that hypoxic and glucose-deprived cancer cells enter a quiescent state supported by mitophagy, fatty acid β-oxidation, and amino acid catabolism, concurrently enhancing their invasive capabilities. Reoxygenation and glucose restoration efficiently reversed cell quiescence without affecting cellular viability, highlighting significant molecular plasticity in adapting to microenvironmental challenges. Furthermore, cancer cells exhibited substantial perturbation of protein O-glycosylation, leading to simplified glycophenotypes with shorter glycosidic chains. Exploiting glycoengineered cell models, we established that immature glycosylation contributes to reduced cell proliferation and increased invasion. Our findings collectively indicate that hypoxia and glucose deprivation trigger cancer aggressiveness, reflecting an adaptive escape mechanism underpinned by altered metabolism and protein glycosylation, providing grounds for clinical intervention.
Keywords: Biological sciences; Cancer; Cancer systems biology; Health sciences; Internal medicine; Medical specialty; Medicine; Natural sciences; Oncology; Systems biology.
© 2025 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Hypoxia enhances the malignant nature of bladder cancer cells and concomitantly antagonizes protein O-glycosylation extension.Oncotarget. 2016 Sep 27;7(39):63138-63157. doi: 10.18632/oncotarget.11257. Oncotarget. 2016. PMID: 27542232 Free PMC article.
-
Glycoproteomics identifies HOMER3 as a potentially targetable biomarker triggered by hypoxia and glucose deprivation in bladder cancer.J Exp Clin Cancer Res. 2021 Jun 9;40(1):191. doi: 10.1186/s13046-021-01988-6. J Exp Clin Cancer Res. 2021. PMID: 34108014 Free PMC article.
-
Prolonged hypoxia switched on cancer stem cell-like plasticity in HepG2 tumourspheres cultured in serum-free media.In Vitro Cell Dev Biol Anim. 2021 Oct;57(9):896-911. doi: 10.1007/s11626-021-00625-y. Epub 2021 Nov 8. In Vitro Cell Dev Biol Anim. 2021. PMID: 34750738
-
Glycobiology Modifications in Intratumoral Hypoxia: The Breathless Side of Glycans Interaction.Cell Physiol Biochem. 2017;41(5):1801-1829. doi: 10.1159/000471912. Epub 2017 Apr 3. Cell Physiol Biochem. 2017. PMID: 28376491 Review.
-
Hypoxia-Induced Cancer Cell Responses Driving Radioresistance of Hypoxic Tumors: Approaches to Targeting and Radiosensitizing.Cancers (Basel). 2021 Mar 4;13(5):1102. doi: 10.3390/cancers13051102. Cancers (Basel). 2021. PMID: 33806538 Free PMC article. Review.
Cited by
-
E-selectin affinity glycoproteomics reveals neuroendocrine proteins and the secretin receptor as a poor-prognosis signature in colorectal cancer.Mol Oncol. 2025 Mar;19(3):635-658. doi: 10.1002/1878-0261.13733. Epub 2024 Nov 7. Mol Oncol. 2025. PMID: 39508360 Free PMC article.
-
Glycomics and Glycoproteomics Reveal Distinct Oligomannose Carriers Across Bladder Cancer Stages.Int J Mol Sci. 2025 May 20;26(10):4891. doi: 10.3390/ijms26104891. Int J Mol Sci. 2025. PMID: 40430030 Free PMC article.
References
LinkOut - more resources
Full Text Sources
