

Functional Characterization of Parallel Fiber-Purkinje Cell Synapses in Two Friedreich's Ataxia Mouse Models
- PMID: 39907933
- PMCID: PMC11799031
- DOI: 10.1007/s12311-025-01796-0
Functional Characterization of Parallel Fiber-Purkinje Cell Synapses in Two Friedreich's Ataxia Mouse Models
Abstract
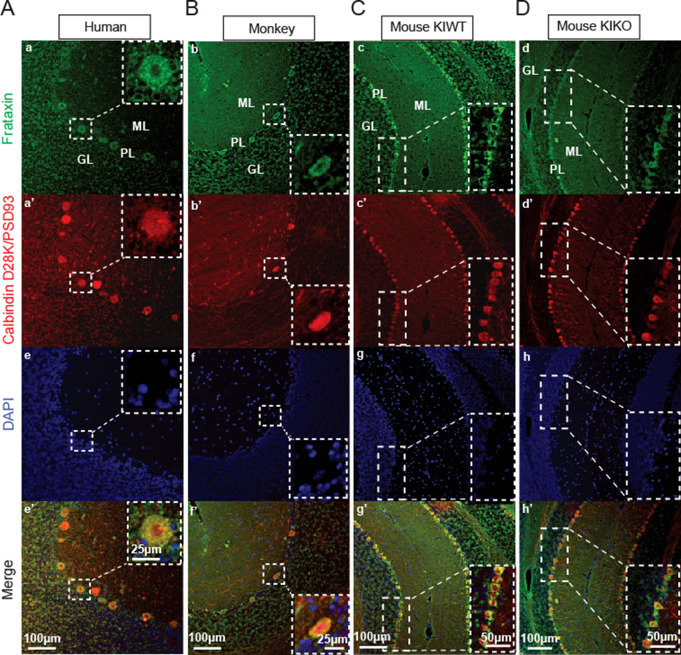
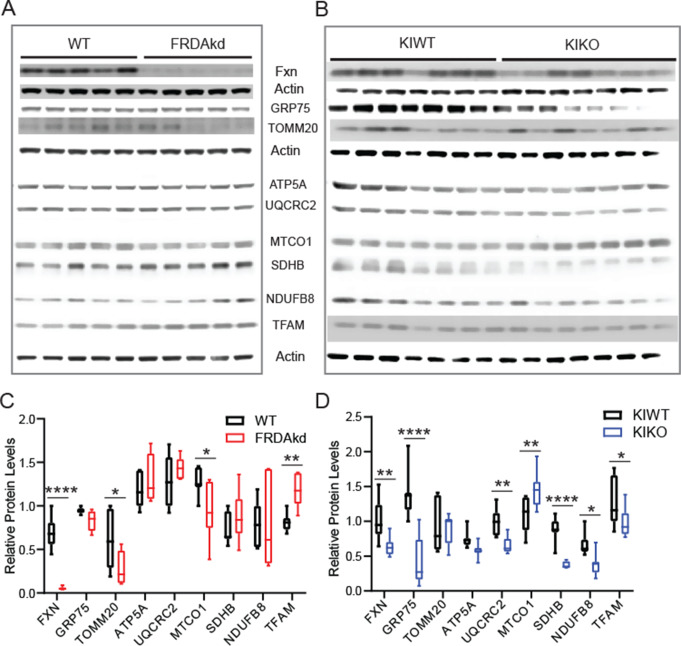
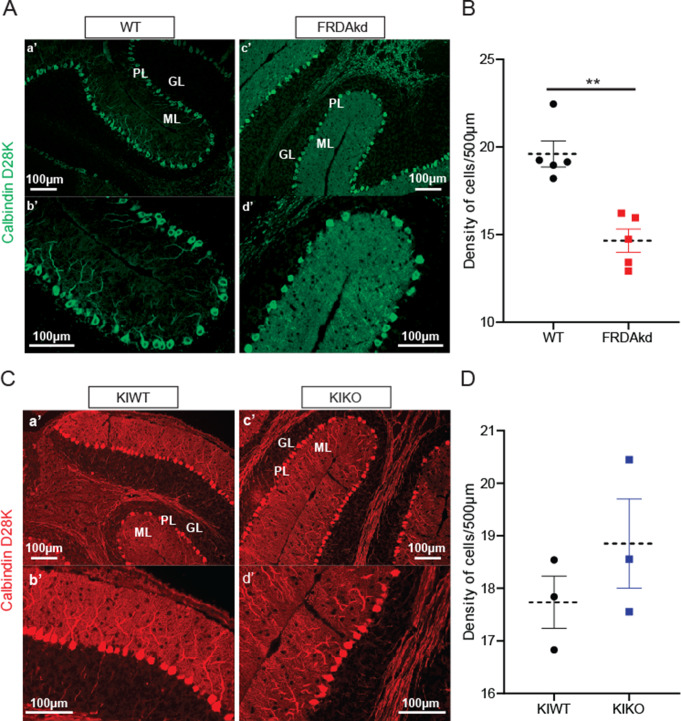
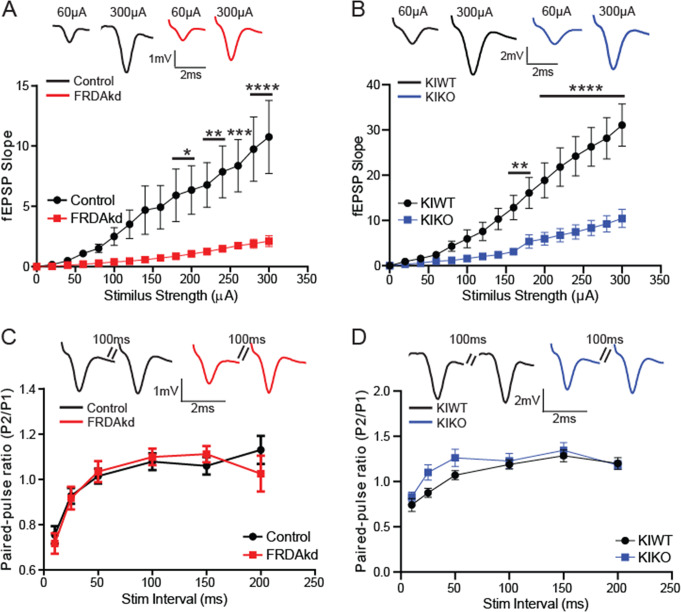
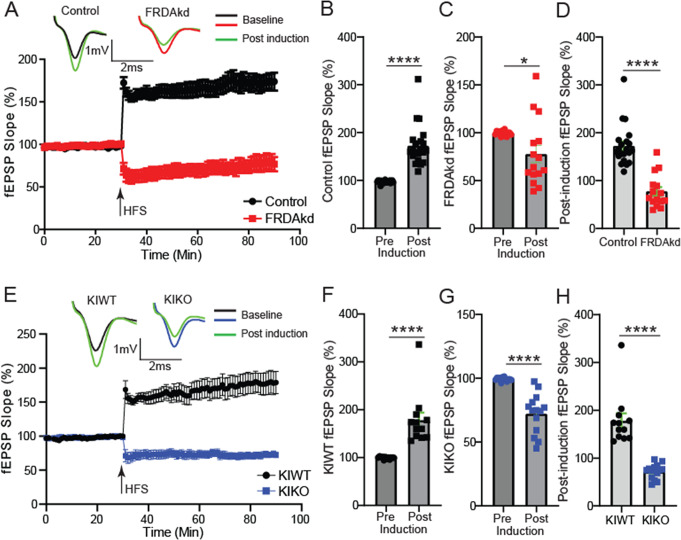
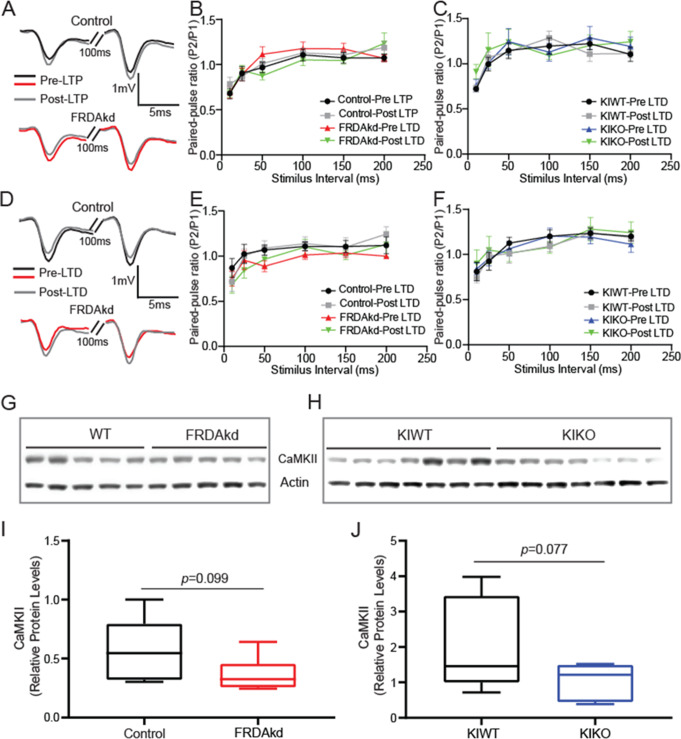
Friedreich ataxia (FRDA) is an autosomal recessive disorder caused by GAA expansions in the FXN gene, which codes for the protein frataxin (FXN). These mutations reduce FXN expression, leading to mitochondrial dysfunction and multisystemic disease. Accumulating evidence suggests that neuronal dysfunction, rather than neuronal death, may drive the neurological phenotypes of FRDA, but the mechanisms underlying such neurological phenotypes remain unclear. To investigate the neural circuit basis of this dysfunction, we employed field recordings to measure Purkinje cell (PC) function and synaptic properties along with western blotting and immunohistochemistry to determine their density and structure in two established FRDA mouse models, the shRNA-frataxin (FRDAkd) and the frataxin knock in-knockout (KIKO) mice. Western blotting demonstrated subtle changes in mitochondrial proteins and only a modest reduction in the density of calbindin positive cells PCs in the cerebellar cortex of the FRDAkd mice, with no change in the density of PCs in the KIKO mice. Though PC density differed slightly in the two models, field recordings of parallel fiber-PC synapses in the molecular layer demonstrated concordant hypo-excitability of basal synaptic transmission and impairments of long-term plasticity using induction protocols associated with both potentiation and depression of synaptic strength. These results indicate that synaptic instability might be a common feature in FRDA mouse models.
Keywords: And Mitochondria; Cerebellum; Frataxin; Friedreich’s ataxia; Long-term plasticity; Synaptic transmission.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Generative AI in Scientific Writing: The authors declare that generative artificial intelligence (AI) and AI-assisted technologies were not used in the writing process of this manuscript. Competing Interests: The authors declare no competing interests.
Figures


References
-
- Campuzano V, Montermini L, Molto MD, Pianese L, Cossee M, Cavalcanti F, Monros E, Rodius F, Duclos F, Monticelli A, Zara F, Canizares J, Koutnikova H, Bidichandani SI, Gellera C, Brice A, Trouillas P, De Michele G, Filla A, De Frutos R, Palau F, Patel PI, Di Donato S, Mandel JL, Cocozza S, Koenig M, Pandolfo M. Friedreich’s ataxia: autosomal recessive disease caused by an intronic GAA triplet repeat expansion. Science. 1996;271:1423–7. 10.1126/science.271.5254.1423. - DOI - PubMed
-
- Pandolfo P M. Friedreich ataxia: the clinical picture. J Neurol. 2009;256(Suppl 1):3–8. 10.1007/s00415-009-1002-3. - PubMed
-
- Campuzano V, Montermini L, Lutz Y, Cova L, Hindelang C, Jiralerspong S, Trottier Y, Kish SJ, Faucheux B, Trouillas P, Authier FJ, Durr A, Mandel JL, Vescovi A, Pandolfo M, Koenig M. Frataxin is reduced in Friedreich ataxia patients and is associated with mitochondrial membranes. Hum Mol Genet. 1997;6:1771–80. 10.1093/hmg/6.11.1771. - DOI - PubMed
-
- Guillon B, Bulteau AL, Wattenhofer-Donze M, Schmucker S, Friguet B, Puccio H, Drapier JC, Bouton C. Frataxin deficiency causes upregulation of mitochondrial lon and ClpP proteases and severe loss of mitochondrial Fe-S proteins. FEBS J. 2009;276:1036–47. 10.1111/j.1742-4658.2008.06847.x. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
