

The genetic origin of the Indo-Europeans
- PMID: 39910300
- PMCID: PMC11922553
- DOI: 10.1038/s41586-024-08531-5
The genetic origin of the Indo-Europeans
Erratum in
-
Author Correction: The genetic origin of the Indo-Europeans.Nature. 2025 Mar;639(8054):E14. doi: 10.1038/s41586-025-08789-3. Nature. 2025. PMID: 39979458 No abstract available.
Abstract
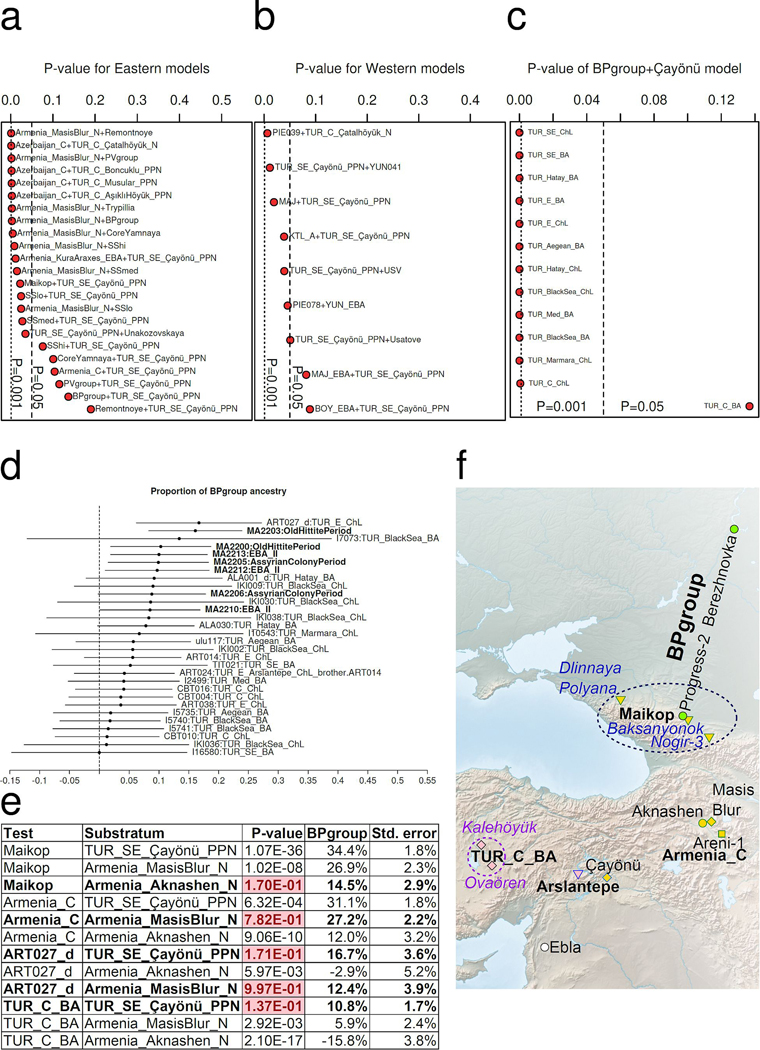
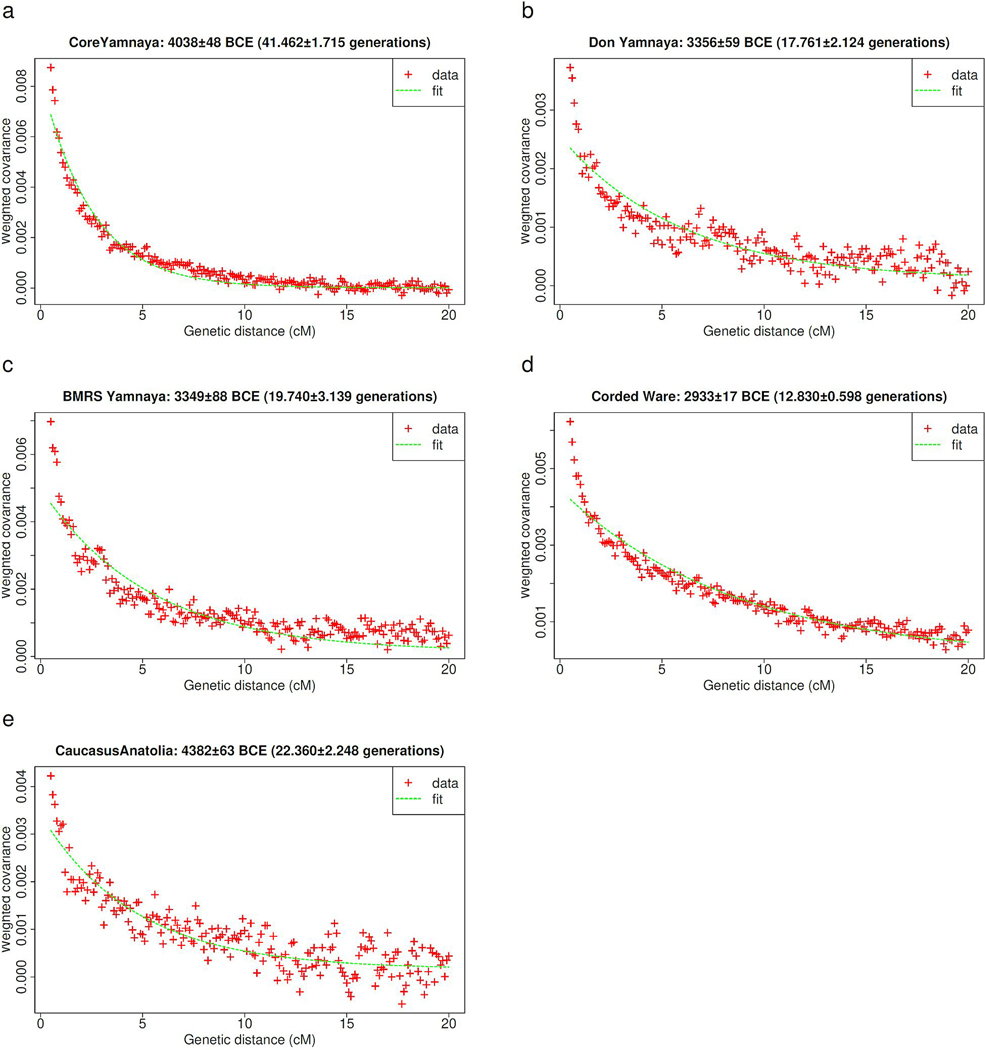
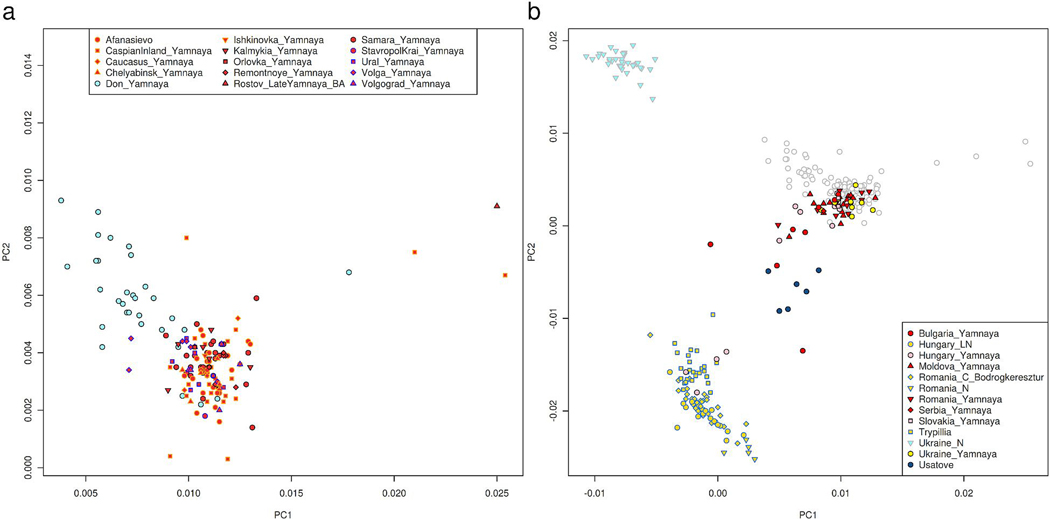
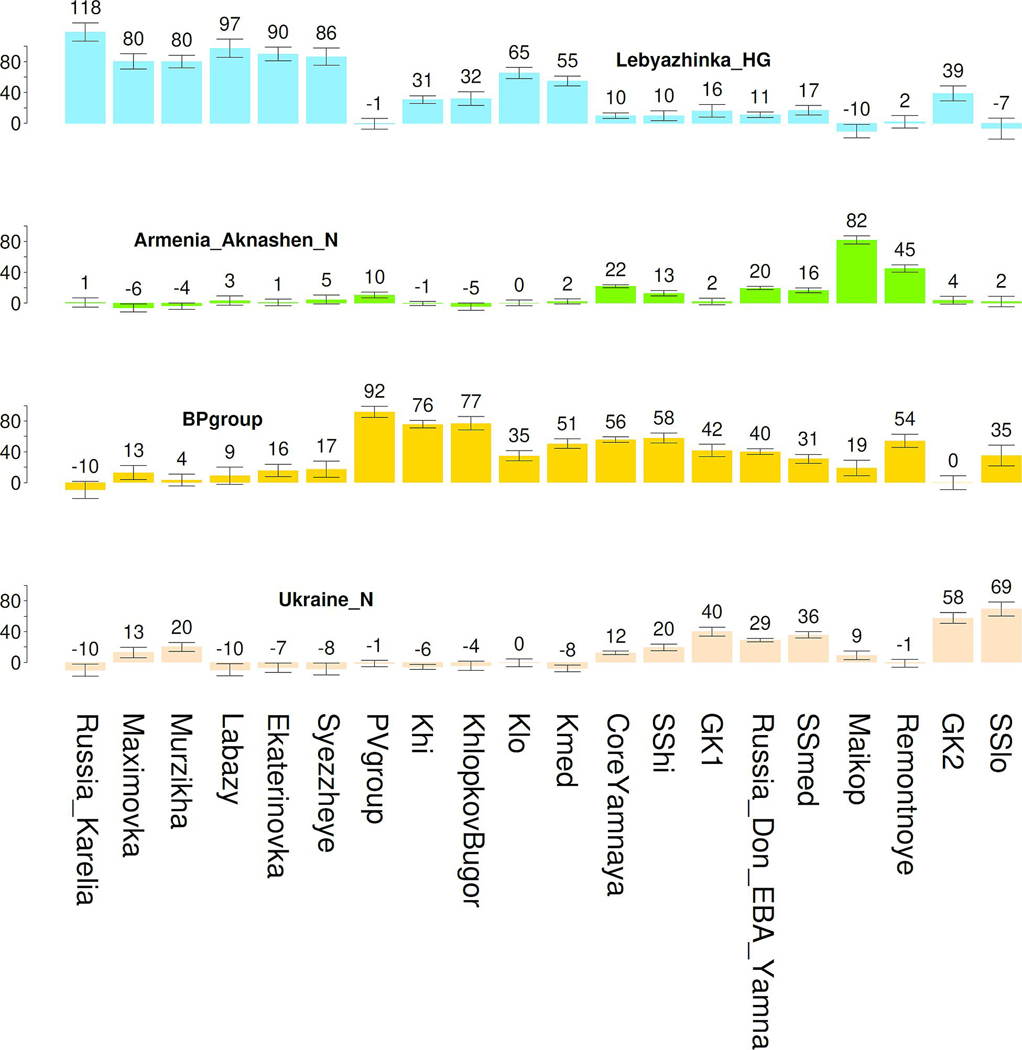
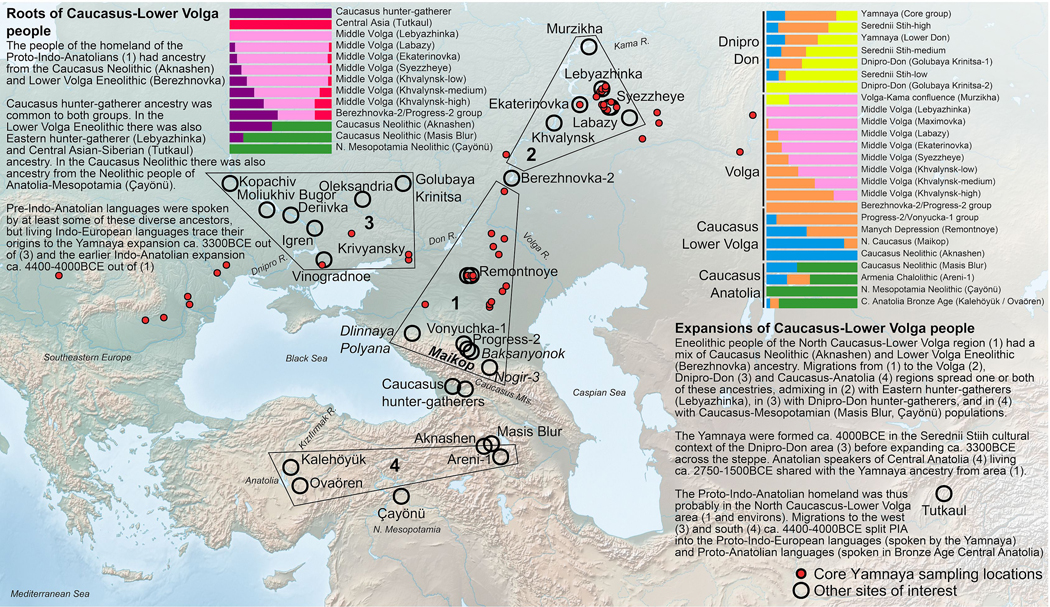
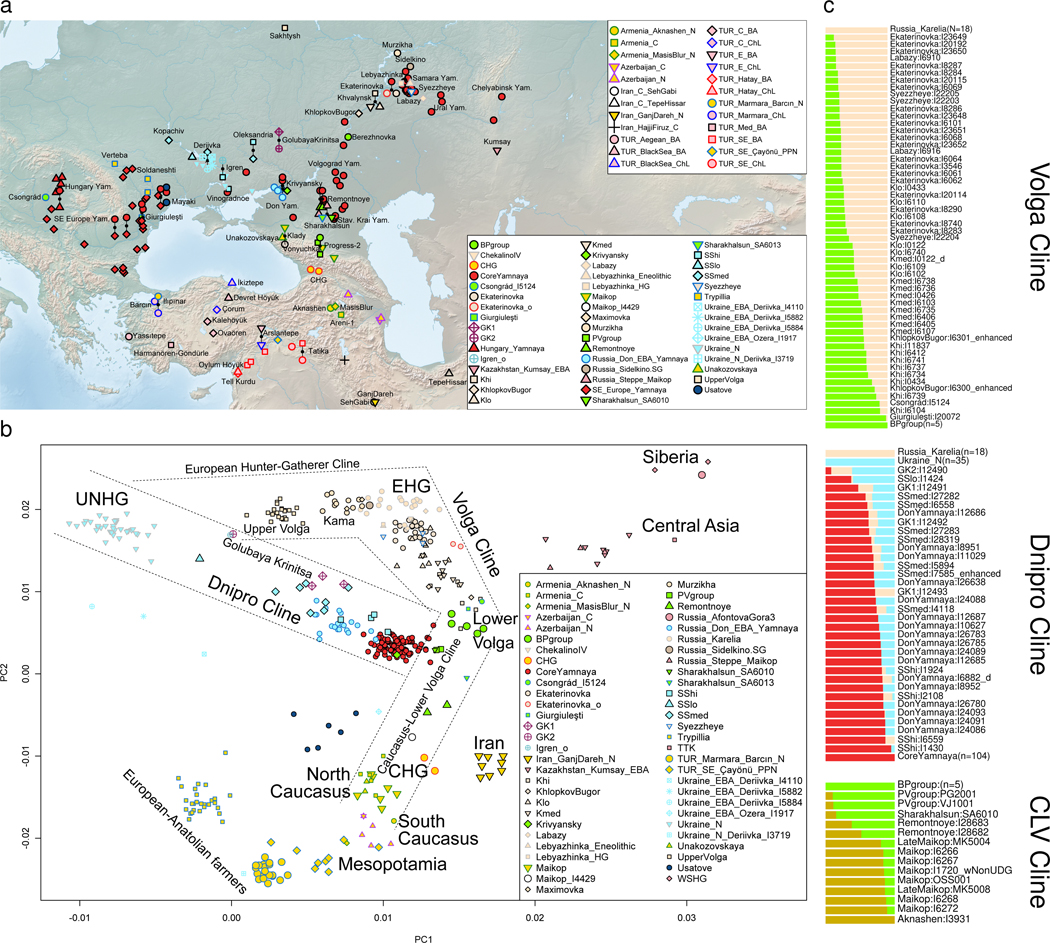
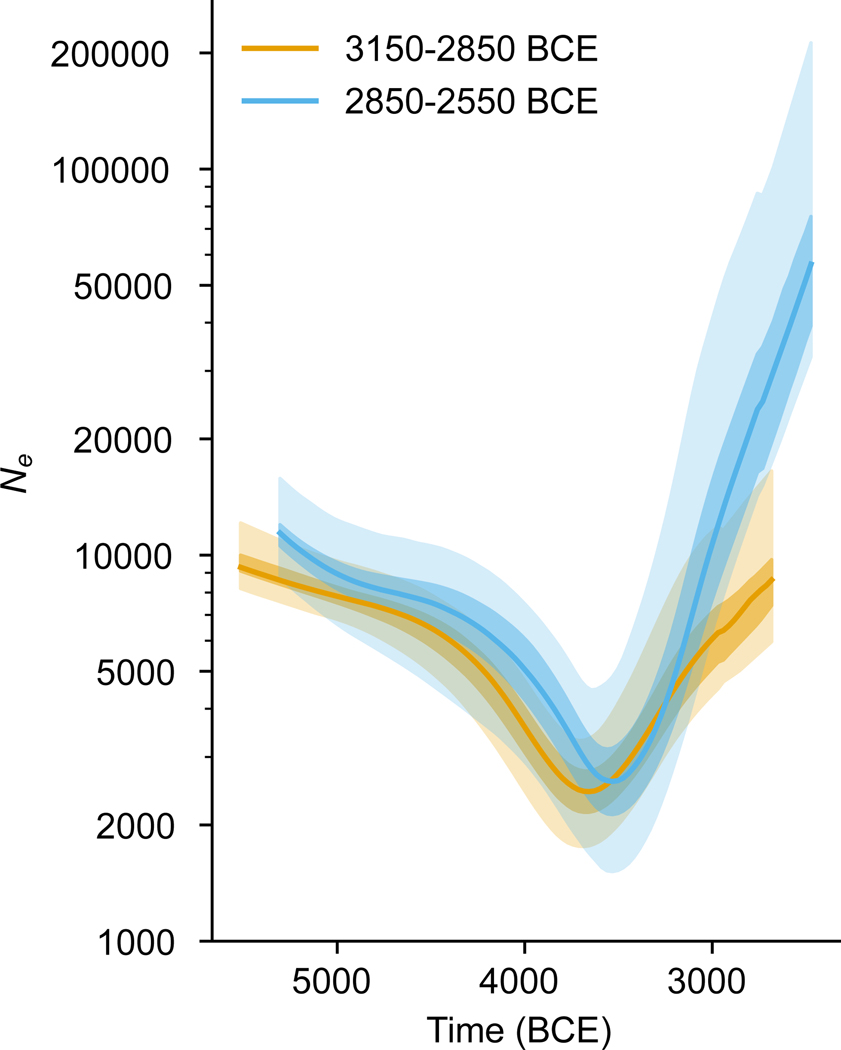
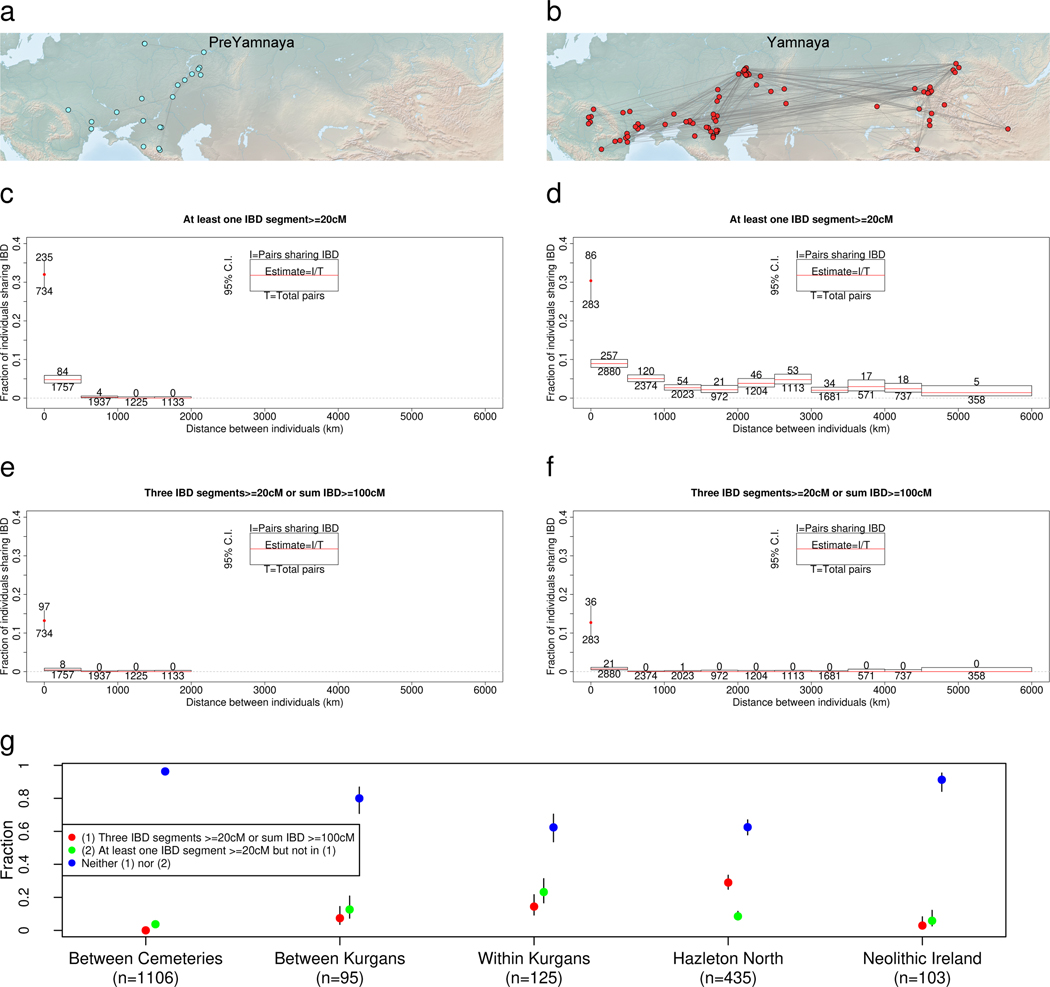
The Yamnaya archaeological complex appeared around 3300 BC across the steppes north of the Black and Caspian Seas, and by 3000 BC it reached its maximal extent, ranging from Hungary in the west to Kazakhstan in the east. To localize Yamnaya origins among the preceding Eneolithic people, we assembled ancient DNA from 435 individuals, demonstrating three genetic clines. A Caucasus-lower Volga (CLV) cline suffused with Caucasus hunter-gatherer1 ancestry extended between a Caucasus Neolithic southern end and a northern end at Berezhnovka along the lower Volga river. Bidirectional gene flow created intermediate populations, such as the north Caucasus Maikop people, and those at Remontnoye on the steppe. The Volga cline was formed as CLV people mixed with upriver populations of Eastern hunter-gatherer2 ancestry, creating hypervariable groups, including one at Khvalynsk. The Dnipro cline was formed when CLV people moved west, mixing with people with Ukraine Neolithic hunter-gatherer ancestry3 along the Dnipro and Don rivers to establish Serednii Stih groups, from whom Yamnaya ancestors formed around 4000 BC and grew rapidly after 3750-3350 BC. The CLV people contributed around four-fifths of the ancestry of the Yamnaya and, entering Anatolia, probably from the east, at least one-tenth of the ancestry of Bronze Age central Anatolians, who spoke Hittite4,5. We therefore propose that the final unity of the speakers of 'proto-Indo-Anatolian', the language ancestral to both Anatolian and Indo-European people, occurred in CLV people some time between 4400 BC and 4000 BC.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


Update of
-
The Genetic Origin of the Indo-Europeans.bioRxiv [Preprint]. 2024 Apr 18:2024.04.17.589597. doi: 10.1101/2024.04.17.589597. bioRxiv. 2024. Update in: Nature. 2025 Mar;639(8053):132-142. doi: 10.1038/s41586-024-08531-5. PMID: 38659893 Free PMC article. Updated. Preprint.
References
-
- Kloekhorst A. in The Indo-European Puzzle Revisited: Integrating Archaeology, Genetics, and Linguistics (eds Eske Willerslev, Guus Kroonen, & Kristian Kristiansen) 42–60 (Cambridge University Press, 2023).
-
- Kroonen G, Barjamovic G. & Peyrot M. Linguistic supplement to Damgaard et al. 2018 : Early Indo-European languages, Anatolian, Tocharian and Indo-Iranian. (2018). <https://zenodo.org/doi/10.5281/zenodo.1240523>. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
