

Limited impact of Salmonella stress and persisters on antibiotic clearance
- PMID: 39910302
- PMCID: PMC11882453
- DOI: 10.1038/s41586-024-08506-6
Limited impact of Salmonella stress and persisters on antibiotic clearance
Abstract
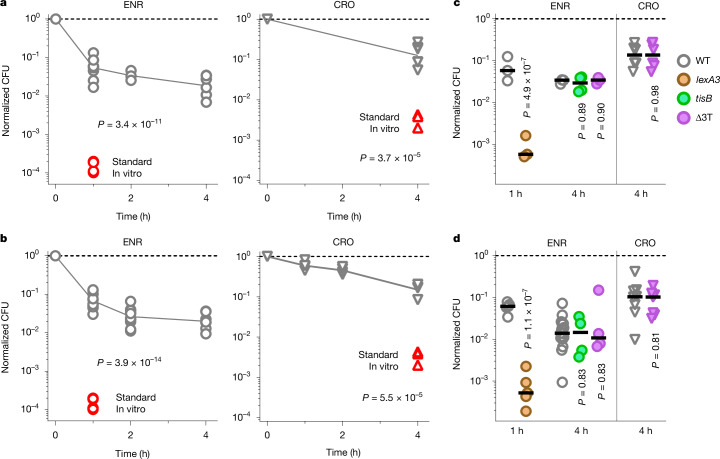
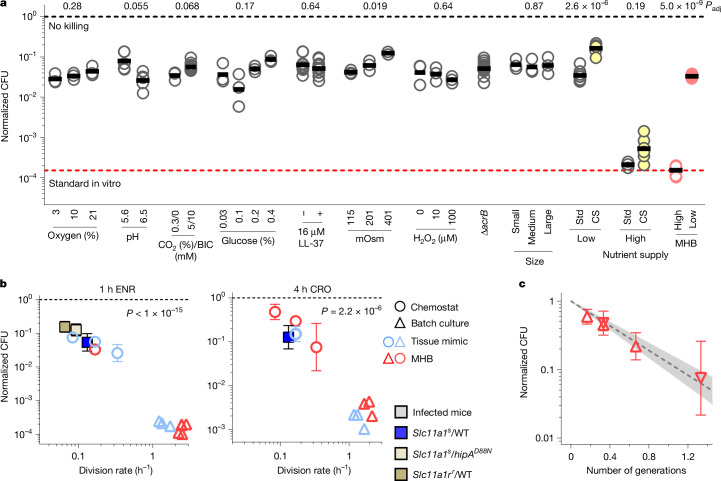
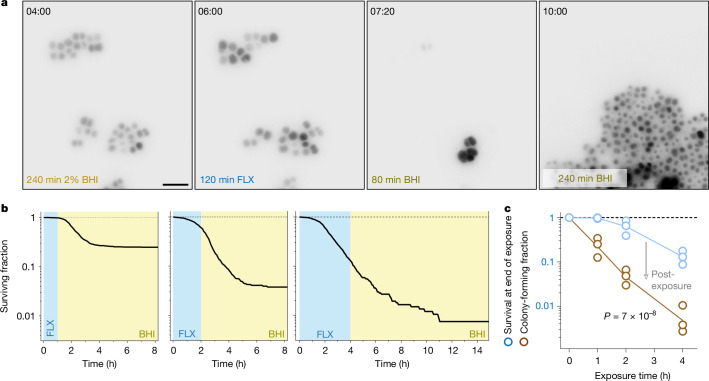
Antimicrobial compounds are essential for controlling bacterial infections. Stress-induced bacterial tolerance and persisters can undermine antimicrobial activities under laboratory conditions, but their quantitative effects under physiological conditions remain unclear1,2. Here we determined constraints on clearance of Salmonella by antimicrobials in infected mice and tissue-mimicking chemostats. The antibiotics enrofloxacin and ceftriaxone exhibited poor anti-Salmonella activity under both conditions, primarily owing to severe nutrient starvation, which restricted Salmonella replication3-5. Other infection-associated conditions, such as acidic pH, glucose, oxidative stress, nitrosative stress, antimicrobial peptides, osmolarity, oxygen limitation, carbon dioxide and carbonate, as well as drug efflux, toxin-antitoxin modules and cell size had limited effects. A subset of resilient Salmonella appeared as a key obstacle for clearance by enrofloxacin, based on the biphasic decline of Salmonella colony-forming units. However, these data were misleading, because colony formation was confounded by extensive post-exposure killing. More accurate single-cell, real-time assays showed uniformly slow damage, indicating high resilience across the entire Salmonella population. The resulting extensive survival of bulk bacteria minimized the effect of hyper-resilient persisters. Thus, starvation-induced general resilience of Salmonella was the main cause of poor antibiotic clearance. These findings highlight the importance of quantifying antibiotic activity with real-time, single-cell assays under physiological conditions.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
-
- Claudi, B. et al. Phenotypic variation of Salmonella in host tissues delays eradication by antimicrobial chemotherapy. Cell158, 722–733 (2014). - PubMed
-
- Cunrath, O. & Bumann, D. Host resistance factor SLC11A1 restricts Salmonella growth through magnesium deprivation. Science366, 995–999 (2019). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
