

New insights into the N-glycomes of Dictyostelium species
- PMID: 39911813
- PMCID: PMC11795075
- DOI: 10.1016/j.bbadva.2025.100142
New insights into the N-glycomes of Dictyostelium species
Abstract
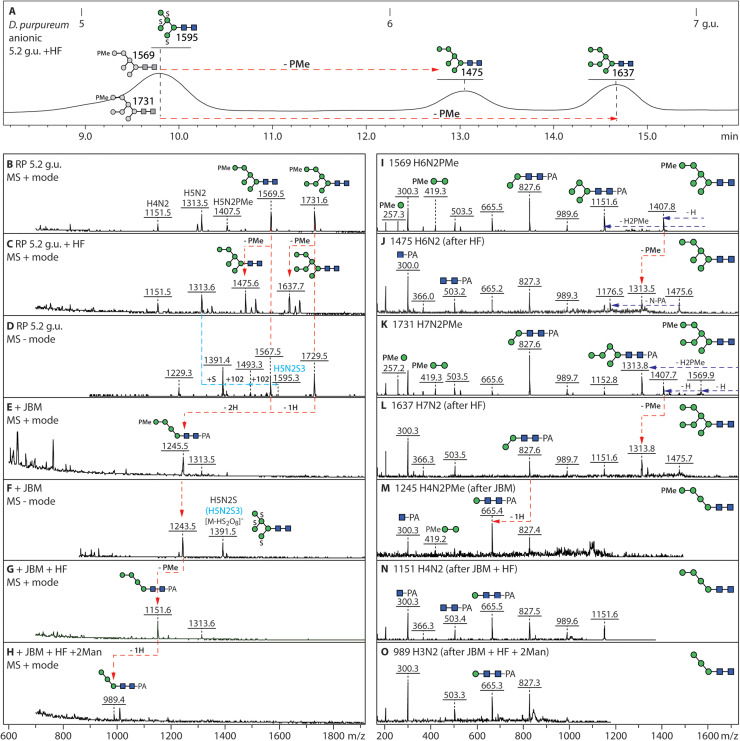
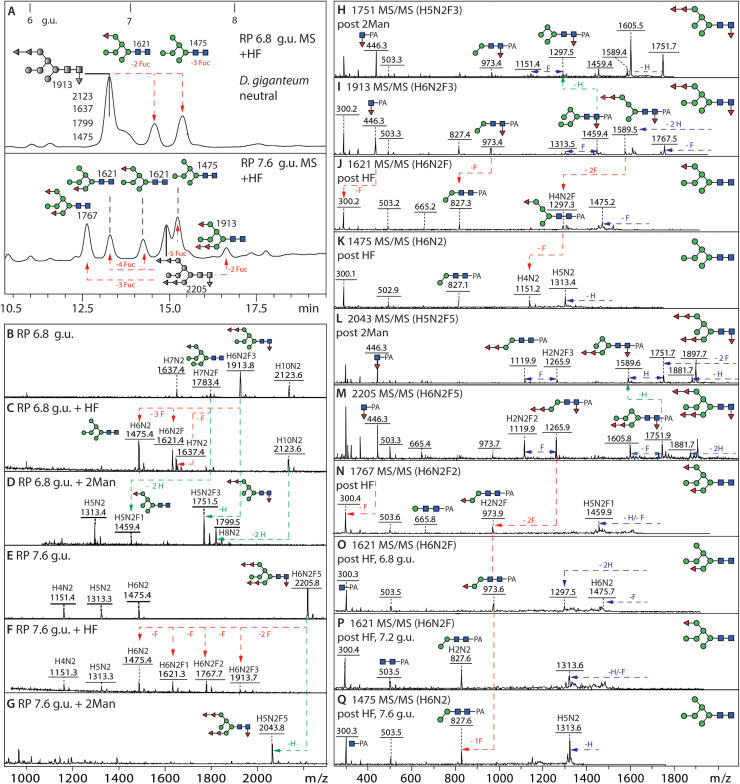
Dictyostelia are cellular slime molds, a group of Amoebozoa, that form multicellular fruiting bodies out of aggregating cells able of differentiating into resistant spore forms. In previous studies on Dictyostelium discoideum, it was demonstrated that their N-glycans, as in most eukaryotes, derive from the Glc3Man9GlcNAc2-PP-Dol precursor; however, unique glyco-epitopes, including intersecting GlcNAc, core α1,3-fucosylation, sulphation and methylphosphorylation, were detected. In the present study, we have examined the N-glycans of two other Dictyostelium species, D. purpureum, whose genome is also sequenced, and D. giganteum. The detailed glycomic analysis of their fruiting bodies was based on isomeric separation of the glycan structures by HPLC, followed by mass spectrometry in combination with enzymatic digests and chemical treatments. Two features absent from the 'model' dictyostelid D. discoideum were found: especially in D. purpureum, a long linear galactose arm β1,4-linked to the β1,4-N-acetylglucosamine on the 'lower' A-branch of its oligo-mannosylated structures could be identified. In contrast, neutral N-glycans with multiple fucose residues attached to terminal mannoses were found in D. giganteum. All three species have common modifications on their anionic N-glycans: while (methyl)phosphorylated residues are always associated with terminal mannose residues, the sulphation position differs. While D. discoideum has 6-sulphation of subterminal mannose residues, D. giganteum and D. purpureum may rather have 2-sulphation of core α1,6-mannose. Overall, we have discovered species-specific glycan variations and our data will contribute to future comparative and functional studies on these three species within the same genus.
Keywords: Dictyostelia; Fucose; Galactose; N-glycan; Phosphate; Sulphate.
© 2025 The Author(s). Published by Elsevier B.V.
Conflict of interest statement
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Katharina Paschinger reports financial support was provided by Austrian Science Fund. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






References
-
- Sheikh S., Thulin M., Cavender J.C., Escalante R., Kawakami S.I., Lado C., Landolt J.C., Nanjundiah V., Queller D.C., Strassmann J.E., Spiegel F.W., Stephenson S.L., Vadell E.M., Baldauf S.L. A new classification of the dictyostelids. Protist. 2018;169(1):1–28. - PubMed
-
- Loomis W.F. Comparative genomics of the dictyostelids. Methods Mol. Biol. 2013;983:39–58. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
