

REDD1 expression in podocytes facilitates renal inflammation and pyroptosis in streptozotocin-induced diabetic nephropathy
- PMID: 39920111
- PMCID: PMC11806006
- DOI: 10.1038/s41419-025-07396-4
REDD1 expression in podocytes facilitates renal inflammation and pyroptosis in streptozotocin-induced diabetic nephropathy
Abstract
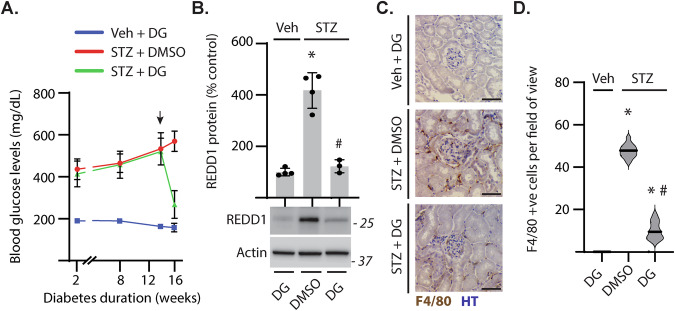
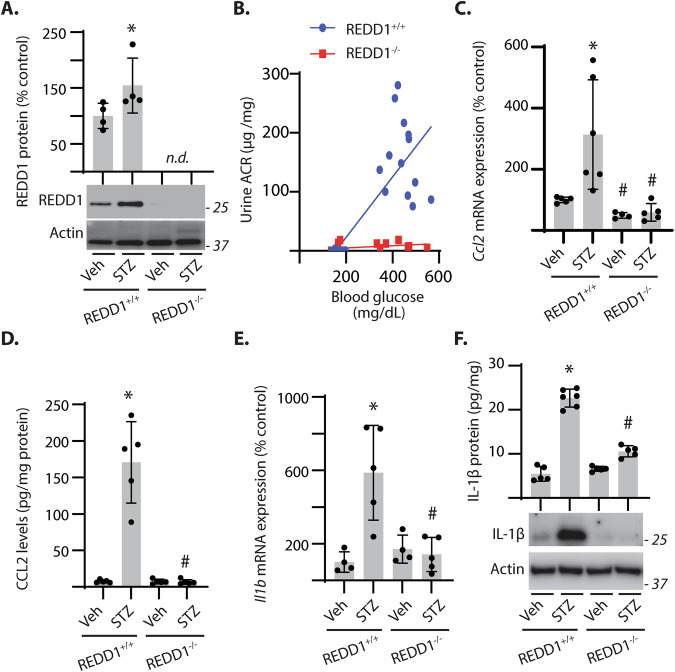
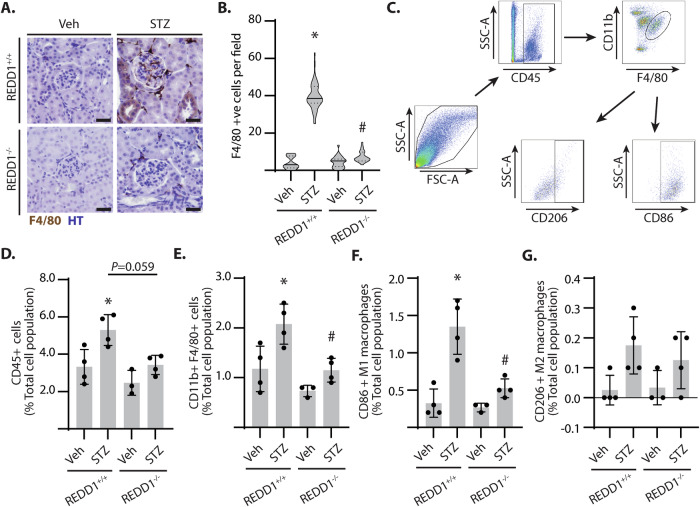
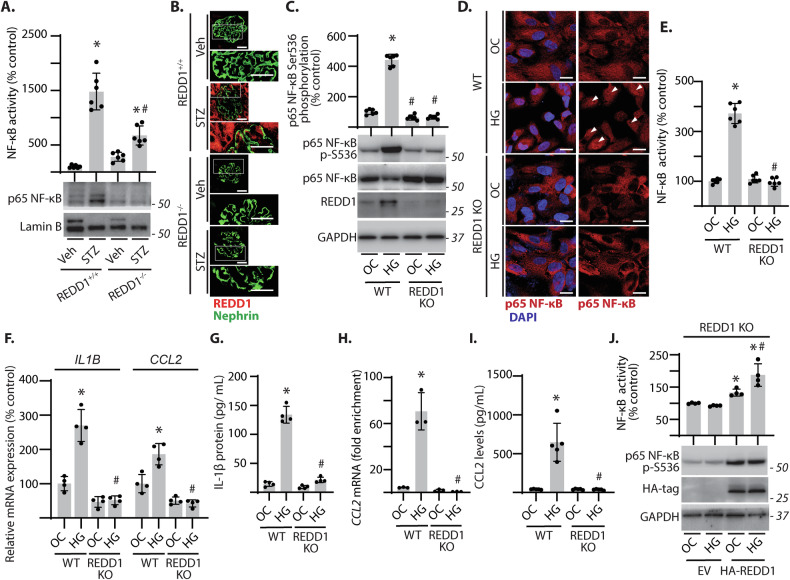
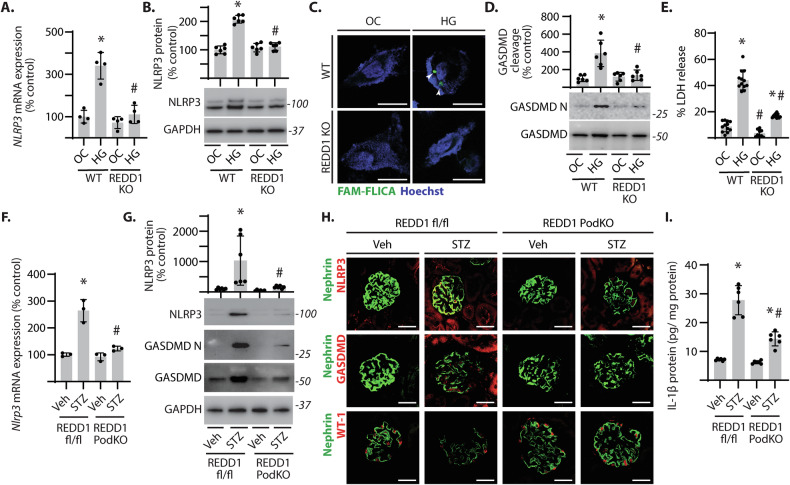
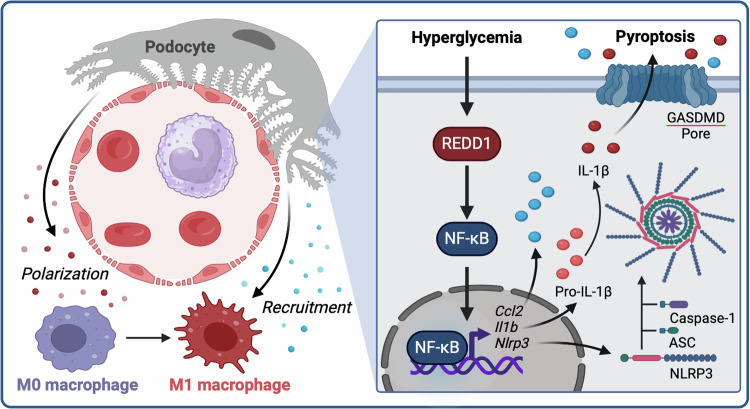
Sterile inflammation resulting in an altered immune response is a key determinant of renal injury in diabetic nephropathy (DN). In this investigation, we evaluated the hypothesis that hyperglycemic conditions augment the pro-inflammatory immune response in the kidney by promoting podocyte-specific expression of the stress response protein regulated in development and DNA damage response 1 (REDD1). In support of the hypothesis, streptozotocin (STZ)-induced diabetes increased REDD1 protein abundance in the kidney concomitant with renal immune cell infiltration. In diabetic mice, administration of the SGLT2 inhibitor dapagliflozin was followed by reductions in blood glucose concentration, renal REDD1 protein abundance, and immune cell infiltration. In contrast with diabetic REDD1+/+ mice, diabetic REDD1-/- mice did not exhibit albuminuria, increased pro-inflammatory factors, or renal macrophage infiltration. In cultured human podocytes, exposure to hyperglycemic conditions promoted REDD1-dependent activation of NF-κB signaling. REDD1 deletion in podocytes attenuated both the increase in chemokine expression and macrophage chemotaxis under hyperglycemic conditions. Notably, podocyte-specific REDD1 deletion prevented the pro-inflammatory immune cell infiltration in the kidneys of diabetic mice. Furthermore, exposure of podocytes to hyperglycemic conditions promoted REDD1-dependent pyroptotic cell death, evidenced by an NLRP3-mediated increase in caspase-1 activity and LDH release. REDD1 expression in podocytes was also required for an increase in pyroptosis markers in the glomeruli of diabetic mice. The data support that podocyte-specific REDD1 is necessary for chronic NF-κB activation in the context of diabetes and raises the prospect that therapies targeting podocyte-specific REDD1 may be helpful in DN.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethical approval: All procedures adhered to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Penn State College of Medicine Institutional Animal Care and Use Committee (PROTO202302452).
Figures

References
-
- Koye DN, Shaw JE, Reid CM, Atkins RC, Reutens AT, Magliano DJ. Incidence of chronic kidney disease among people with diabetes: a systematic review of observational studies. Diabet Med J Br Diabet Assoc. 2017;34:887–901. - PubMed
-
- United States Renal Data System. 2022 USRDS Annual Data Report: Epidemiology of kidney disease in the United States. National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD %8, 2022. https://adr.usrds.org/2022.
-
- Xu J, Murphy SL, Kochanek KD, Arias E. Mortality in the United States, 2021. 2022.https://stacks.cdc.gov/view/cdc/122516. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
