

Aspirin inhibits proteasomal degradation and promotes α-synuclein aggregate clearance through K63 ubiquitination
- PMID: 39920137
- PMCID: PMC11806099
- DOI: 10.1038/s41467-025-56737-6
Aspirin inhibits proteasomal degradation and promotes α-synuclein aggregate clearance through K63 ubiquitination
Abstract
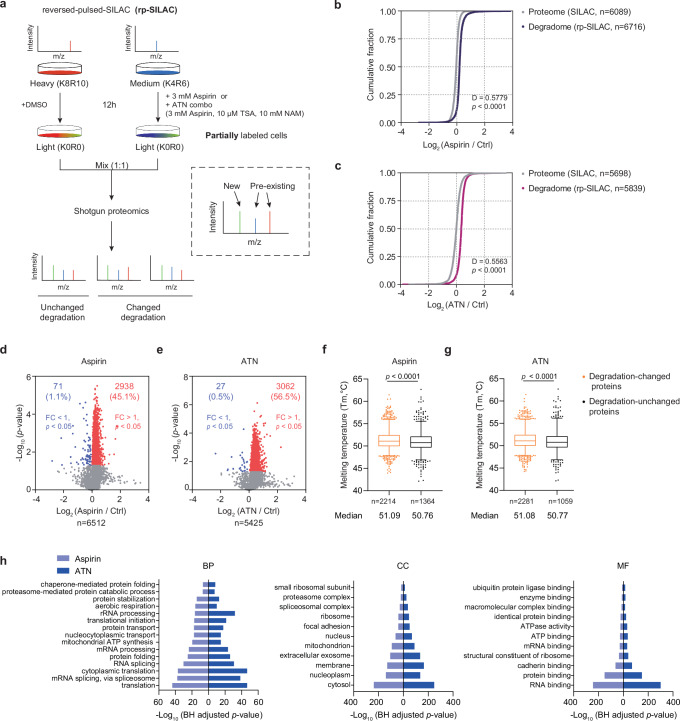
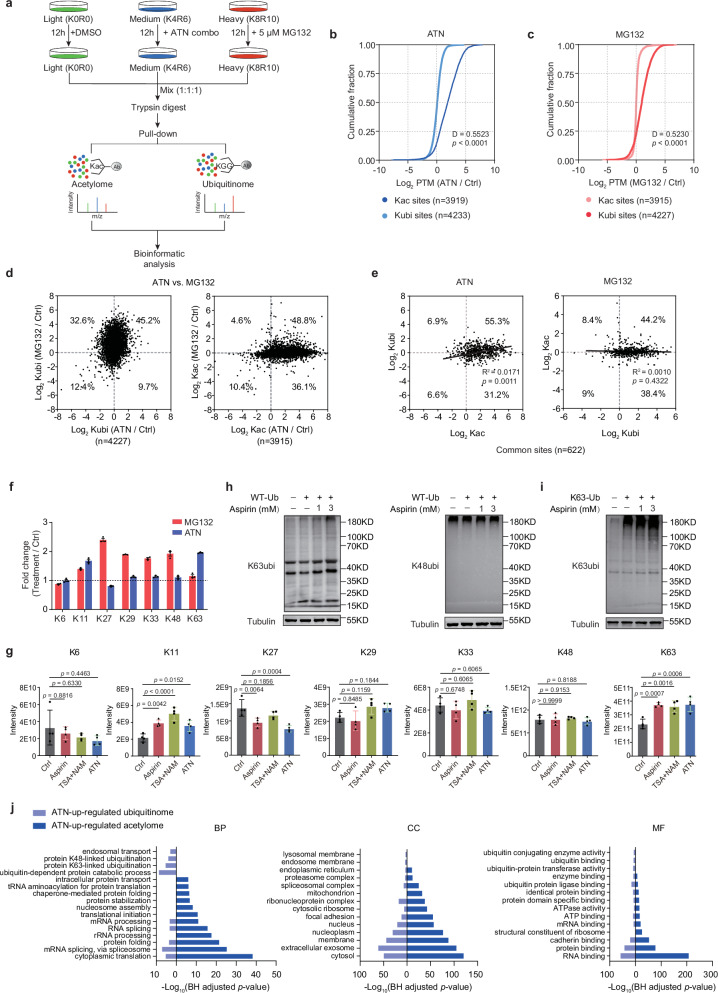
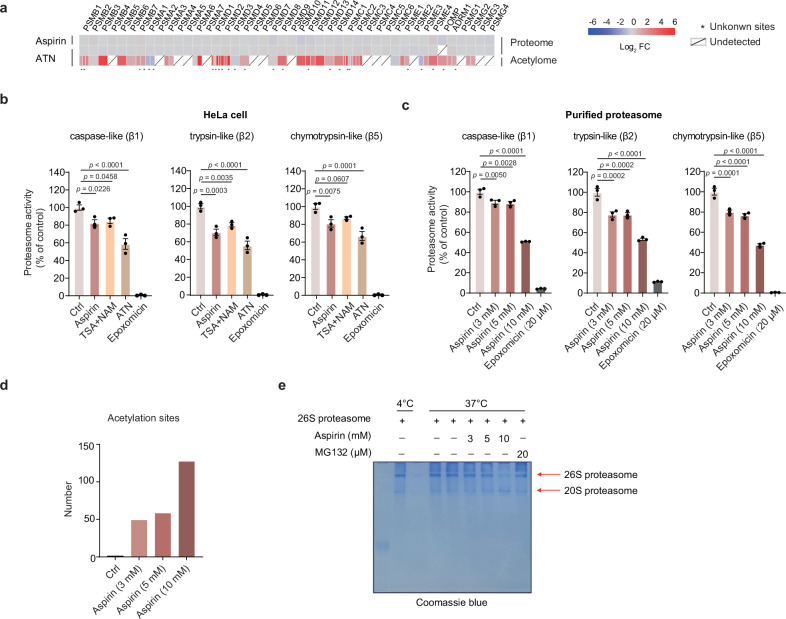
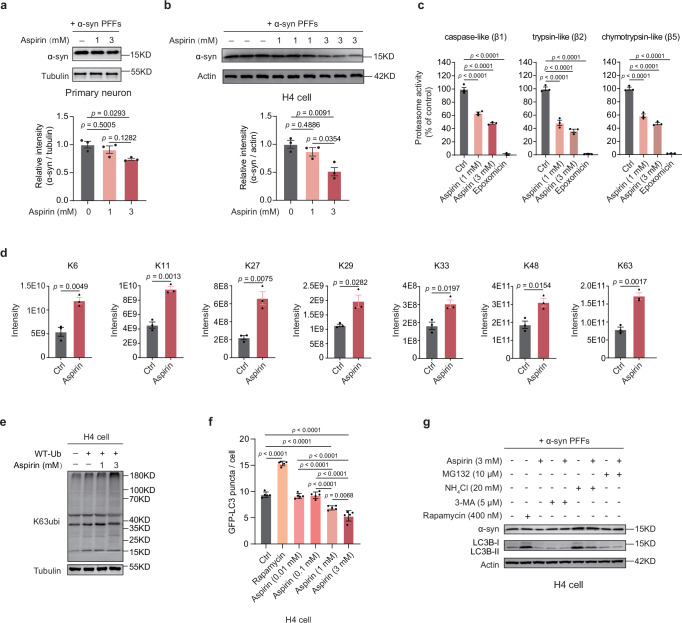
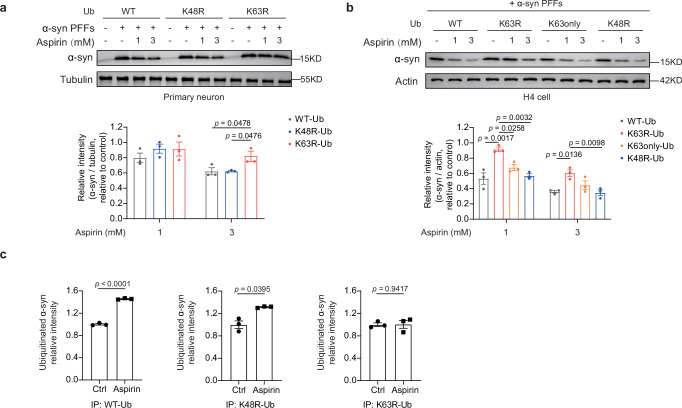
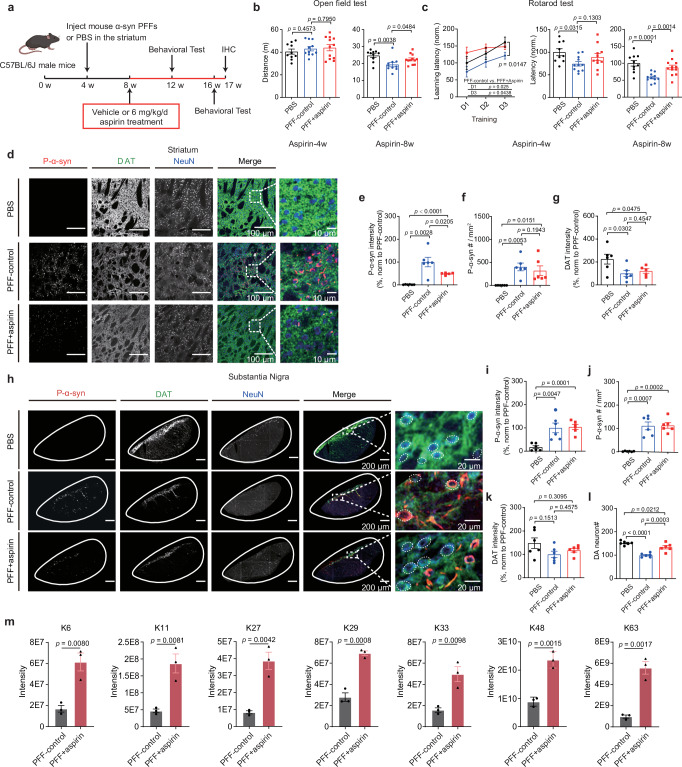
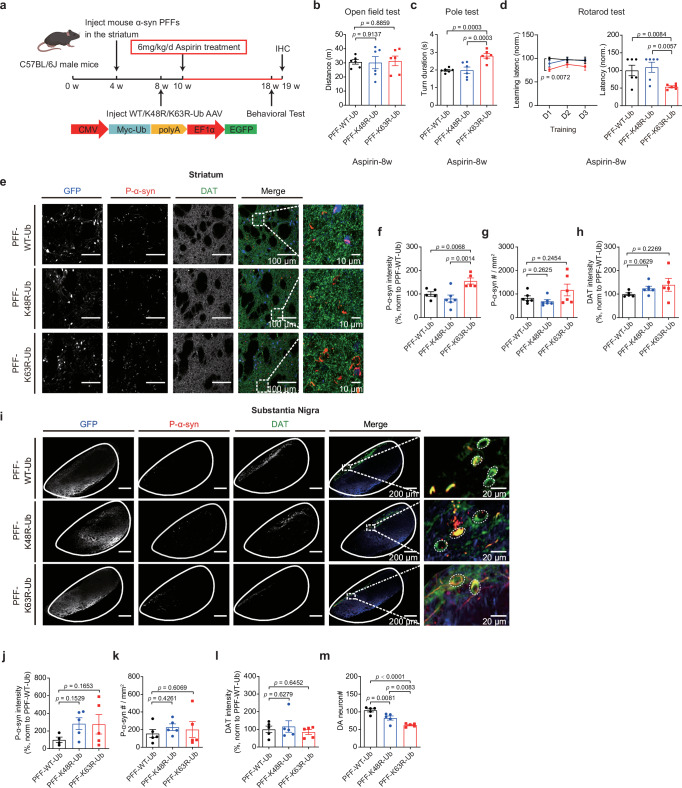
Aspirin is a potent lysine acetylation inducer, but its impact on lysine ubiquitination and ubiquitination-directed protein degradation is unclear. Herein, we develop the reversed-pulsed-SILAC strategy to systematically profile protein degradome in response to aspirin. By integrating degradome, acetylome, and ubiquitinome analyses, we show that aspirin impairs proteasome activity to inhibit proteasomal degradation, rather than directly suppressing lysine ubiquitination. Interestingly, aspirin increases lysosomal degradation-implicated K63-linked ubiquitination. Accordingly, using the major pathological protein of Parkinson's disease (PD), α-synuclein (α-syn), as an example of protein aggregates, we find that aspirin is able to reduce α-syn in cultured cells, neurons, and PD model mice with rescued locomotor ability. We further reveal that the α-syn aggregate clearance induced by aspirin is K63-ubiquitination dependent in both cells and PD mice. These findings suggest two complementary mechanisms by which aspirin regulates the degradation of soluble and insoluble proteins, providing insights into its diverse pharmacological effects that can aid in future drug development efforts.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: Y.Z., J.G. and K.H. have filed a patent application related to the use of aspirin or its derivatives for treating diseases associated with abnormal aggregation of α-syn. All other authors declare no competing interests. Ethics: All animal experimental procedures were reviewed and approved by the Institutional Animal Care and Use Committee at the Interdisciplinary Research Center on Biology and Chemistry, Chinese Academy of Sciences, and are in accordance with the Guide for the Care and Use of Laboratory Animals of the Chinese Academy of Sciences.
Figures
Similar articles
-
The role of ubiquitin linkages on alpha-synuclein induced-toxicity in a Drosophila model of Parkinson's disease.J Neurochem. 2009 Jul;110(1):208-19. doi: 10.1111/j.1471-4159.2009.06124.x. Epub 2009 Apr 27. J Neurochem. 2009. PMID: 19457126
-
The role of polo-like kinases 2 in the proteasomal and lysosomal degradation of alpha-synuclein in neurons.FASEB J. 2024 Oct;38(20):e70121. doi: 10.1096/fj.202401035R. FASEB J. 2024. PMID: 39436202 Free PMC article.
-
Neuronal Bmi-1 is critical for melatonin induced ubiquitination and proteasomal degradation of α-synuclein in experimental Parkinson's disease models.Neuropharmacology. 2021 Aug 15;194:108372. doi: 10.1016/j.neuropharm.2020.108372. Epub 2020 Nov 4. Neuropharmacology. 2021. PMID: 33157086
-
α-Synuclein ubiquitination and novel therapeutic targets for Parkinson's disease.CNS Neurol Disord Drug Targets. 2014;13(4):630-7. doi: 10.2174/18715273113126660195. CNS Neurol Disord Drug Targets. 2014. PMID: 24168368 Review.
-
Atypical Ubiquitination and Parkinson's Disease.Int J Mol Sci. 2022 Mar 28;23(7):3705. doi: 10.3390/ijms23073705. Int J Mol Sci. 2022. PMID: 35409068 Free PMC article. Review.
Cited by
-
Ubiquitination regulation of mitochondrial homeostasis: a new sight for the treatment of gastrointestinal tumors.Front Immunol. 2025 Mar 11;16:1533007. doi: 10.3389/fimmu.2025.1533007. eCollection 2025. Front Immunol. 2025. PMID: 40134432 Free PMC article. Review.
-
Aspirin Attenuates Liver Fibrosis via Autophagy Induction.J Cell Mol Med. 2025 Jul;29(13):e70696. doi: 10.1111/jcmm.70696. J Cell Mol Med. 2025. PMID: 40611443 Free PMC article.
-
Protein Misfolding and Aggregation as a Mechanistic Link Between Chronic Pain and Neurodegenerative Diseases.Curr Issues Mol Biol. 2025 Apr 8;47(4):259. doi: 10.3390/cimb47040259. Curr Issues Mol Biol. 2025. PMID: 40699658 Free PMC article. Review.
References
-
- Prasad, K. N., Wu, M. & Bondy, S. C. Telomere shortening during aging: Attenuation by antioxidants and anti-inflammatory agents. Mech. Ageing Dev.164, 61–66 (2017). - PubMed
-
- Flossmann, E. & Rothwell, P. M. Effect of aspirin on long-term risk of colorectal cancer: consistent evidence from randomised and observational studies. Lancet369, 1603–1613 (2007). - PubMed
-
- Ayyadevara, S. et al. Aspirin inhibits oxidant stress, reduces age-associated functional declines, and extends lifespan of Caenorhabditis elegans. Antioxid. Redox Signal18, 481–490 (2013). - PubMed
-
- Green, G. A. Understanding NSAIDs: from aspirin to COX-2. Clin. Cornerstone3, 50–60 (2001). - PubMed
-
- Guo, L., Gao, J., Gao, Y., Zhu, Z. & Zhang, Y. Aspirin reshapes acetylomes in inflammatory and cancer cells via CoA-dependent and CoA-independent pathways. J. Proteome19, 962–972 (2020). - PubMed
MeSH terms
Substances
Grants and funding
- 32071245/National Natural Science Foundation of China (National Science Foundation of China)
- 20ZR1474400/Natural Science Foundation of Shanghai (Natural Science Foundation of Shanghai Municipality)
- 2019SHZDZX02/Science and Technology Commission of Shanghai Municipality (Shanghai Municipal Science and Technology Commission)
LinkOut - more resources
Full Text Sources
Miscellaneous
