

Prodomain processing controls BMP-10 bioactivity and targeting to fibrillin-1 in latent conformation
- PMID: 39921464
- PMCID: PMC11806408
- DOI: 10.1096/fj.202401694R
Prodomain processing controls BMP-10 bioactivity and targeting to fibrillin-1 in latent conformation
Abstract
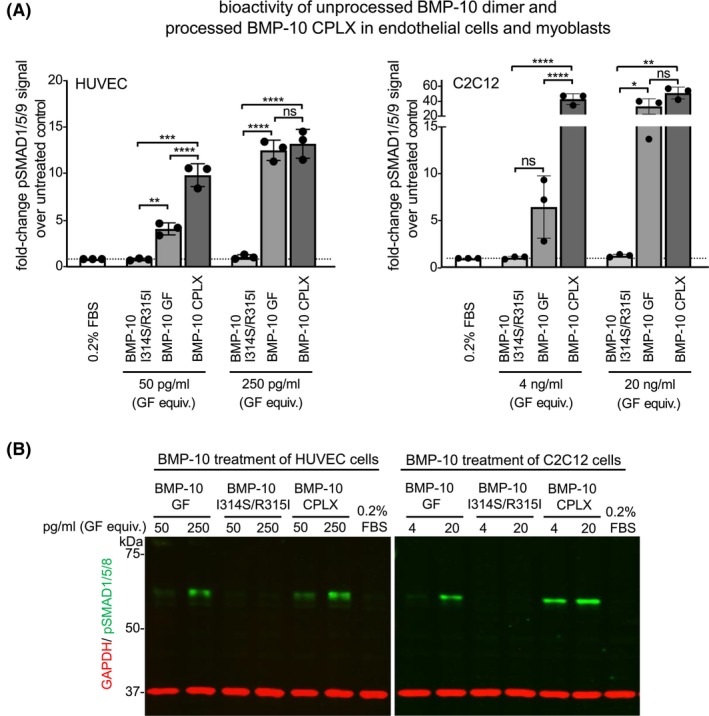
Bone morphogenetic protein 10 (BMP-10) is crucial for endothelial cell signaling via activin receptor-like kinase 1 (ALK1), a pathway central to vascular homeostasis and angiogenesis. Dysregulated BMP-10 signaling contributes to cardiovascular diseases and cancer, highlighting the need to control ALK1-mediated endothelial responses to BMP-10 for therapeutic development. BMP-10 biosynthesis involves processing by proprotein convertases (PPCs) resulting in a non-covalently associated prodomain-growth factor (PD-GF) complex (CPLX), similar to other TGF-β superfamily ligands. However, the molecular requirements for BMP-10 bioactivity remain unclear. We investigated how PPC processing impacts BMP-10 structure, bioactivity, and its interaction with the extracellular matrix (ECM) protein fibrillin-1. Molecular dynamics simulations post-in silico cleavage of the BMP-10 dimer model as well as negative staining and transmission electron microscopy (TEM) revealed that PD processing increases BMP-10 flexibility converting it from a latent wide-angle conformation to a bioactive CPLX which can adopt a V-shape with tighter angle. Only processed BMP-10 demonstrated high potency in HUVEC and C2C12 cells and robust binding to immobilized BMP receptors. Circular dichroism and interaction studies revealed that the N-terminal region of the BMP-10 PD is rich in alpha-helical content, which is essential for efficient complexation with the BMP-10 GF. Binding studies and TEM analyses showed that only the processed BMP-10 CPLX interacts with the N-terminal region of fibrillin-1, causing a conformational change that renders it into a closed ring-shaped conformation. These findings suggest that PD processing induces specific folding events at the PD-GF interface, which is critical for BMP-10 bioactivity and its targeting to the ECM.
Keywords: bone morphogenetic protein; complex; conformational change; electron microscopy; furin; growth factor; molecular dynamics; molecular modeling; proprotein convertases (PPCs); single particle analysis.
© 2025 The Author(s). The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Figures










References
-
- Shi Y, Massague J. Mechanisms of TGF‐beta signaling from cell membrane to the nucleus. Cell. 2003;113:685‐700. - PubMed
-
- Gregory KE, Ono RN, Charbonneau NL, et al. The prodomain of BMP‐7 targets the BMP‐7 complex to the extracellular matrix. J Biol Chem. 2005;280:27970‐27980. - PubMed
-
- Brown MA, Zhao Q, Baker KA, et al. Crystal structure of BMP‐9 and functional interactions with pro‐region and receptors. J Biol Chem. 2005;280:25111‐25118. - PubMed
MeSH terms
Substances
Grants and funding
- U01 HL146188/HL/NHLBI NIH HHS/United States
- 1R56AG075690-01A1/HHS | National Institutes of Health (NIH)
- R01-GM129325/HHS | National Institutes of Health (NIH)
- P41 GM103311/GM/NIGMS NIH HHS/United States
- DFG-INST 216/1174-1/Deutsche Forschungsgemeinschaft (DFG)
- 1R01AG084715/HHS | National Institutes of Health (NIH)
- R01 AG084715/AG/NIA NIH HHS/United States
- R56 AG075690/AG/NIA NIH HHS/United States
- 384170921/Deutsche Forschungsgemeinschaft (DFG)
- ACI-1548562 (83)/National Science Foundation (NSF)
- 203128/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- CMMI 2145759/National Science Foundation (NSF)
- 5U01HL146188/HHS | National Institutes of Health (NIH)
- 468236352/Deutsche Forschungsgemeinschaft (DFG)
- R01 HL133009/HL/NHLBI NIH HHS/United States
- BB/T017643/1/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
- 397484323/Deutsche Forschungsgemeinschaft (DFG)
- BB/S015779/1/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
- R01 GM129325/GM/NIGMS NIH HHS/United States
- P41-GM103311/HHS | National Institutes of Health (NIH)
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
