

TEFM facilitates transition from RNA synthesis to DNA synthesis at H-strand replication origin of mtDNA
- PMID: 39922921
- PMCID: PMC11807126
- DOI: 10.1038/s42003-025-07645-4
TEFM facilitates transition from RNA synthesis to DNA synthesis at H-strand replication origin of mtDNA
Abstract
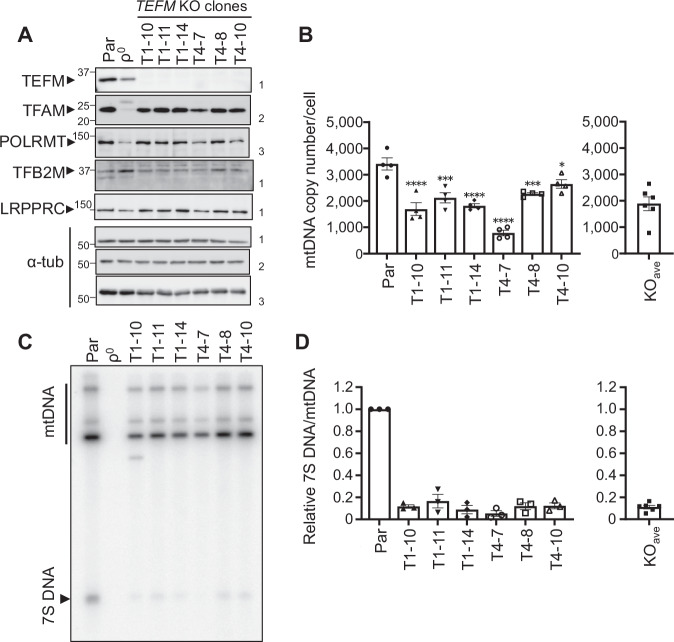
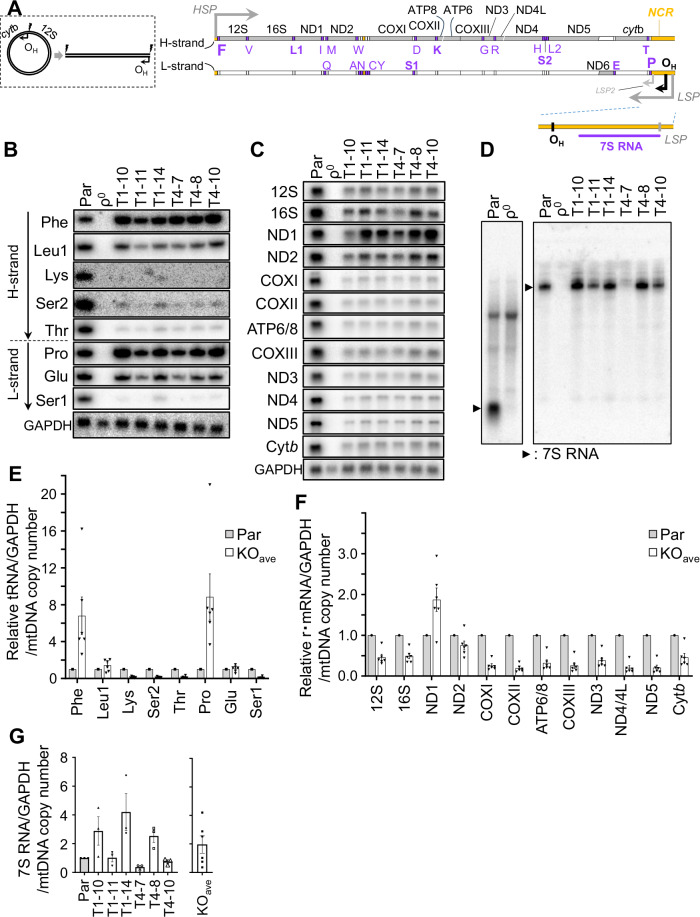
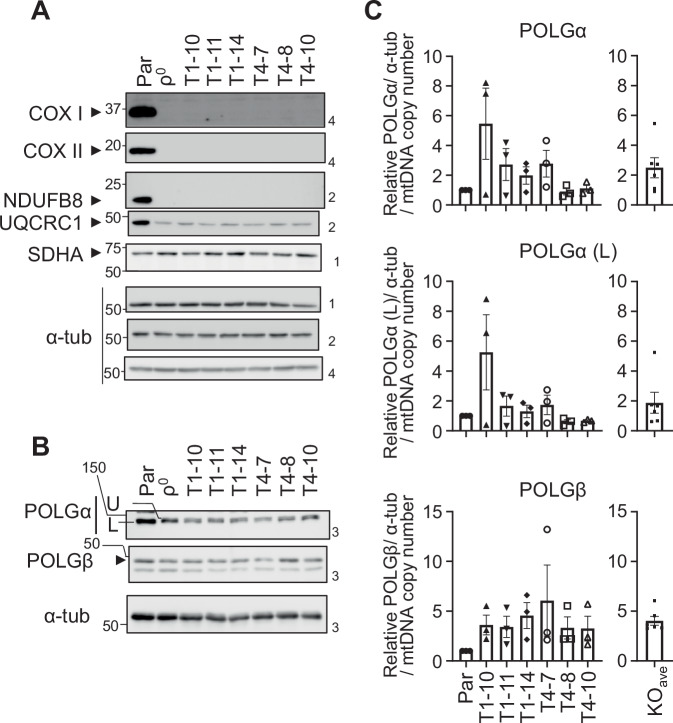
Transcription of human mitochondrial DNA (mtDNA) begins from specific transcription promoters. In strand-asynchronous mtDNA replication, transcripts from the light-strand promoter serve as primers for leading-strand synthesis at the origin of the H-strand replication (OH). A 7S DNA strand, a presumed aborted replication product, is also synthesized from OH. Transition from RNA synthesis to DNA synthesis at OH is crucial for balancing replication with transcription, yet the mechanism remains unclear. Herein, we examine the role of mitochondrial transcription elongation factor (TEFM) in this process. TEFM knockout results in decreased 7S DNA, strand-asynchronous replication intermediates, and mtDNA copy number, all of which are concordant with downregulation of RNA-to-DNA transition at OH. Conversely, levels of tRNAs encoded near transcription promoters increase, indicating enhanced transcription initiation frequency. Taken together, we propose that, in addition to conferring processivity to the mitochondrial RNA polymerase, TEFM plays a crucial role in maintaining the balance between mitochondrial transcription and replication.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare the following competing interests: T.Y. receives research funding through his institution from Renee Medical Corporation. The other authors declare no competing interests.
Figures





References
-
- Anderson, S. et al. Sequence and organization of the human mitochondrial genome. Nature290, 457–465 (1981). - PubMed
-
- Magnusson, J., Orth, M., Lestienne, P. & Taanman, J. W. Replication of mitochondrial DNA occurs throughout the mitochondria of cultured human cells. Exp. Cell. Res.289, 133–142 (2003). - PubMed
-
- Ylikallio, E., Tyynismaa, H., Tsutsui, H., Ide, T. & Suomalainen, A. High mitochondrial DNA copy number has detrimental effects in mice. Hum. Mol. Genet.19, 2695–2705 (2010). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
