

Regulation of lung progenitor plasticity and repair by fatty acid oxidation
- PMID: 39927460
- PMCID: PMC11948574
- DOI: 10.1172/jci.insight.165837
Regulation of lung progenitor plasticity and repair by fatty acid oxidation
Abstract
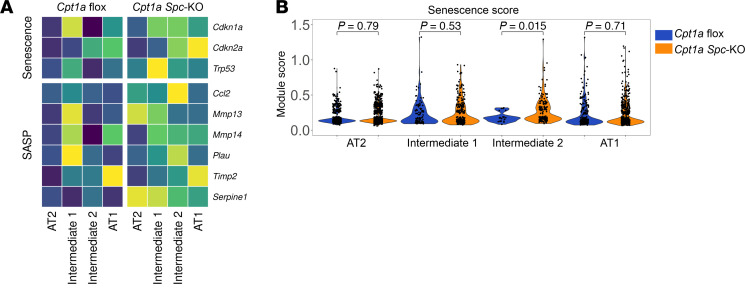
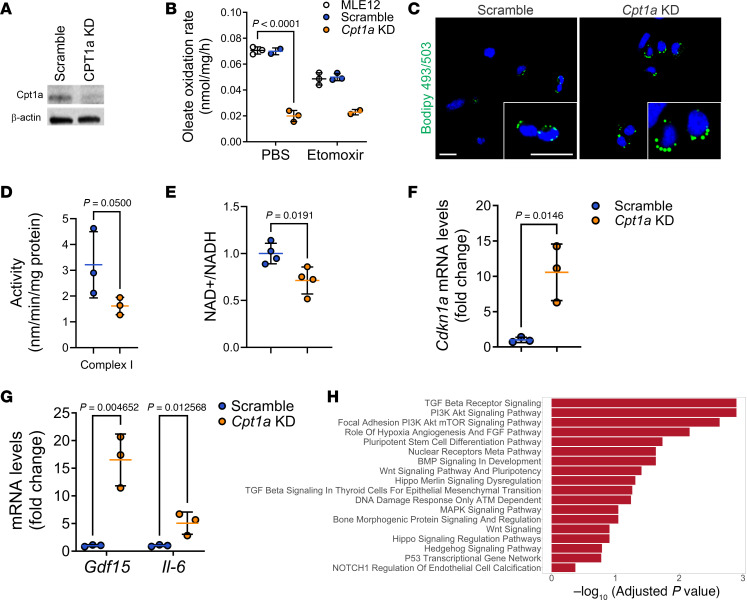
Idiopathic pulmonary fibrosis (IPF) is an age-related interstitial lung disease, characterized by inadequate alveolar regeneration and ectopic bronchiolization. While some molecular pathways regulating lung progenitor cells have been described, the role of metabolic pathways in alveolar regeneration is poorly understood. We report that expression of fatty acid oxidation (FAO) genes is significantly diminished in alveolar epithelial cells of IPF lungs by single-cell RNA sequencing and tissue staining. Genetic and pharmacological inhibition in AT2 cells of carnitine palmitoyltransferase 1a (CPT1a), the rate-limiting enzyme of FAO, promoted mitochondrial dysfunction and acquisition of aberrant intermediate states expressing basaloid, and airway secretory cell markers SCGB1A1 and SCGB3A2. Furthermore, mice with deficiency of CPT1a in AT2 cells show enhanced susceptibility to developing lung fibrosis with an accumulation of epithelial cells expressing markers of intermediate cells, airway secretory cells, and senescence. We found that deficiency of CPT1a causes a decrease in SMAD7 protein levels and TGF-β signaling pathway activation. These findings suggest that the mitochondrial FAO metabolic pathway contributes to the regulation of lung progenitor cell repair responses and deficiency of FAO contributes to aberrant lung repair and the development of lung fibrosis.
Keywords: Fatty acid oxidation; Fibrosis; Metabolism; Mitochondria; Pulmonology.
Conflict of interest statement
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
