

Dynamic Activation of NADPH Oxidases in Immune Responses Modulates Differentiation, Function, and Lifespan of Plasma Cells
- PMID: 39931760
- PMCID: PMC11811814
- DOI: 10.1002/eji.202350975
Dynamic Activation of NADPH Oxidases in Immune Responses Modulates Differentiation, Function, and Lifespan of Plasma Cells
Abstract
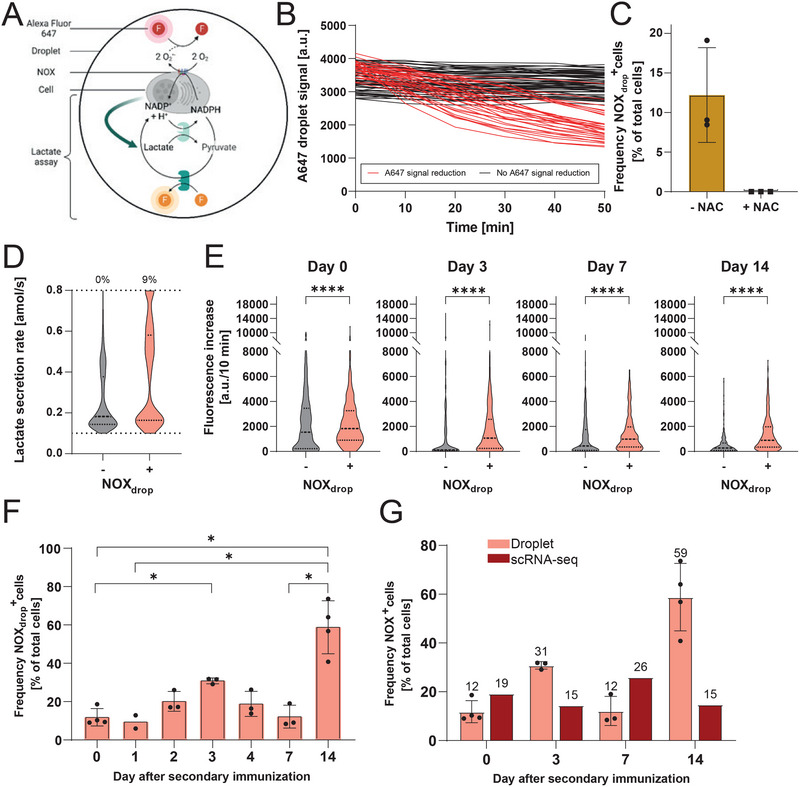
NADPH-oxidase (NOX)-derived reactive oxygen species (ROS) have been described to play essential roles in B-cell activation processes. However, several key questions concerning NOX activity and subsequent ROS production remain unaddressed, including fundamental processes such as differentiation, functional competence, cellular metabolism, and viability. This study investigated these questions in a murine B-cell response after secondary immunization. We combined single-cell transcriptomics and single-cell detection of NOX activity and observed that various subsets of B cells dynamically express NOX1 and NOX2. The NOX+ cellular phenotype correlated with increased activity of metabolic pathways, augmented lactate production, lower IgG secretion rates, and markers for longevity. The NOX+ cellular phenotype was also associated with increased cellular stress and apoptosis, underscoring the intricate relationship between ROS and cellular survival. Consequently, these insights advance our understanding of how long-lived humoral immunity is formed.
Keywords: B cells; BCR signaling; NADPH oxidase; TLR4 signaling; immune metabolism; immune response.
© 2025 The Author(s). European Journal of Immunology published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
