

Bone marrow mesenchymal stromal cells support translation in refractory acute myeloid leukemia
- PMID: 39932190
- PMCID: PMC7617453
- DOI: 10.1016/j.celrep.2024.115151
Bone marrow mesenchymal stromal cells support translation in refractory acute myeloid leukemia
Abstract
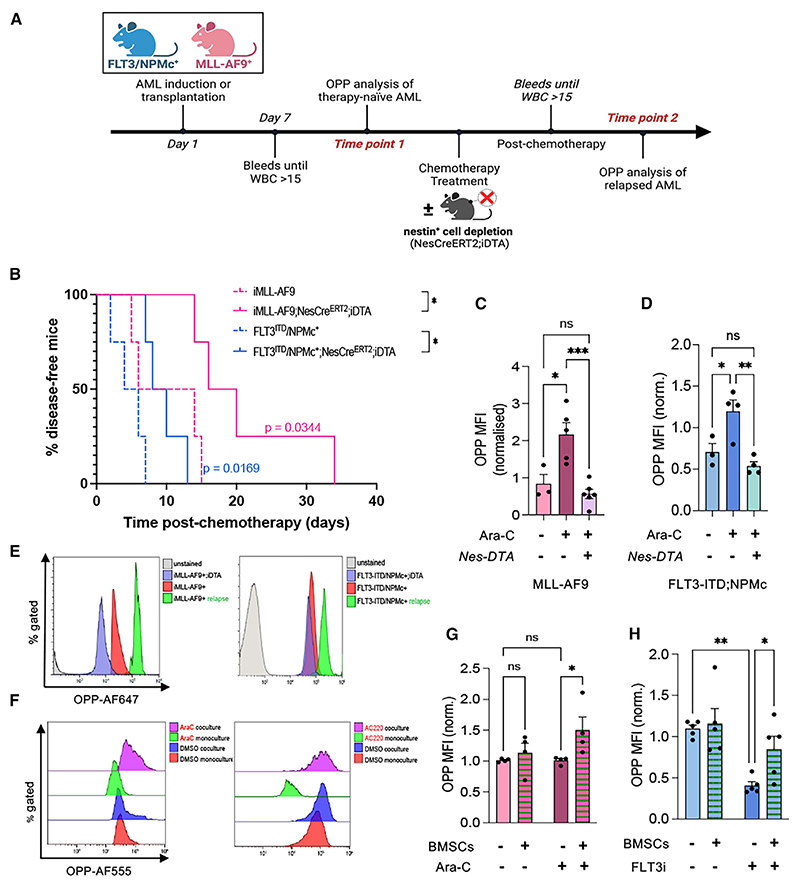
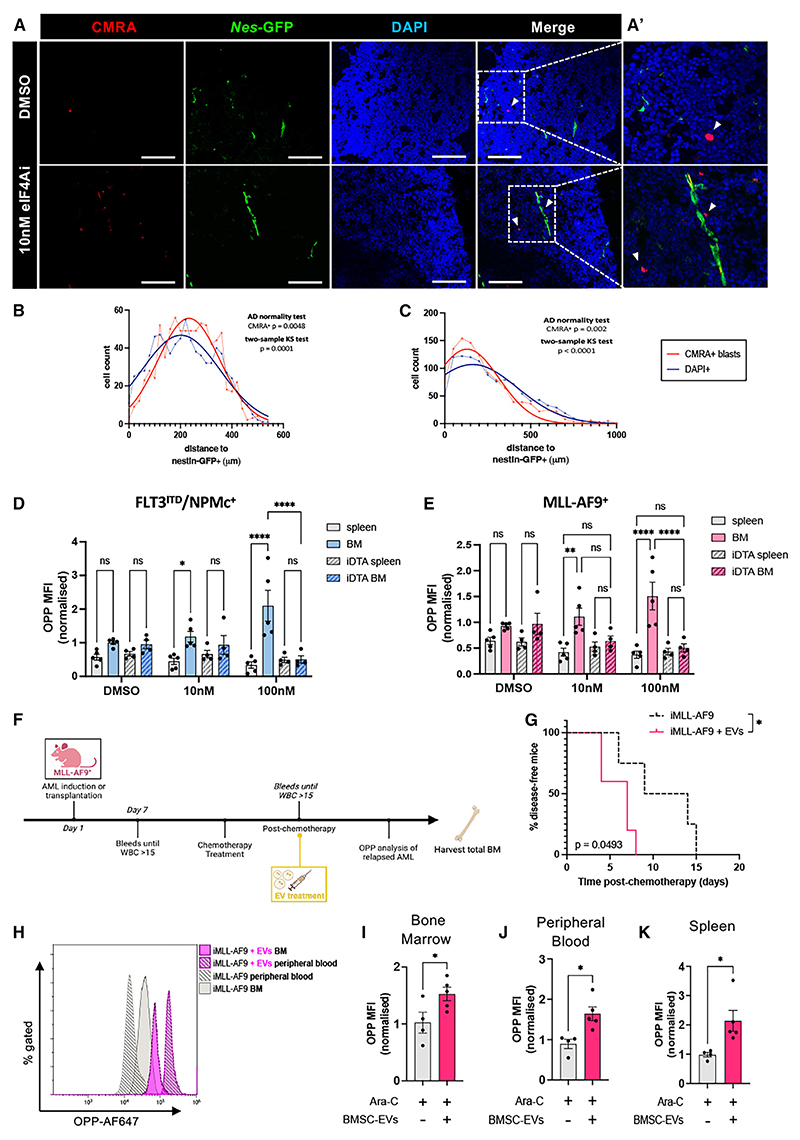
In acute myeloid leukemia (AML), malignant cells surviving chemotherapy rely on high mRNA translation and their microenvironmental metabolic support to drive relapse. However, the role of translational reprogramming in the niche is unclear. Here, we found that relapsing AML cells increase translation in their bone marrow (BM) niches, where BM mesenchymal stromal cells (BMSCs) become a source of eIF4A-cap-dependent translation machinery that is transferred to AML cells via extracellular vesicles (EVs) to meet their translational demands. In two independent models of highly chemo-resistant AML driven by MLL-AF9 or FLT3-ITD (internal tandem duplication) and nucleophosmin (NPMc) mutations, protein synthesis levels increase in refractory AML dependent on nestin+ BMSCs. Inhibiting cap-dependent translation in BMSCs abolishes their chemoprotective ability, while EVs from BMSCs carrying eIF4A boost AML cell translation and survival. Consequently, eIF4A inhibition synergizes with conventional chemotherapy. Together, these results suggest that AML cells rely on BMSCs to maintain an oncogenic translational program required for relapse.
Keywords: CP: Cancer; acute myeloid leukemia; bone marrow mesenchymal stromal cells; chemotherapy; extracellular vesicles; microenvironment; niche; protein synthesis; refractory; relapse; translation.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
-
- Dohner H, Wei AH, Appelbaum FR, Craddock C, DiNardo CD, Dombret H, Ebert BL, Fenaux P, Godley LA, Hasserjian RP, et al. Diagnosis and management of AML in adults: 2022 recommendations from an international expert panel on behalf of the ELN. Blood. 2022;140:1345–1377. - PubMed
-
- Krivtsov AV, Armstrong SA. MLL translocations, histone modifications and leukaemia stem-cell development. Nat Rev Cancer. 2007;7:823–833. - PubMed
-
- Falini B, Nicoletti I, Martelli MF, Mecucci C. Acute myeloid leukemia carrying cytoplasmic/mutated nucleophosmin (NPMc+ AML): biologic and clinical features. Blood. 2007;109:874–885. - PubMed
-
- Forte D, Garcia-Fernandez M, Sanchez-Aguilera A, Stavropoulou V, Fielding C, Martin-Perez D, Lopez JA, Costa ASH, Tronci L, Nikitopoulou E, et al. Bone Marrow Mesenchymal Stem Cells Support Acute Myeloid Leukemia Bioenergetics and Enhance Antioxidant Defense and Escape from Chemotherapy. Cell Metabol. 2020;32:829–843. doi: 10.1016/j.cmet.2020.09.001. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
