

Human and mouse proteomics reveals the shared pathways in Alzheimer's disease and delayed protein turnover in the amyloidome
- PMID: 39934151
- PMCID: PMC11814087
- DOI: 10.1038/s41467-025-56853-3
Human and mouse proteomics reveals the shared pathways in Alzheimer's disease and delayed protein turnover in the amyloidome
Abstract
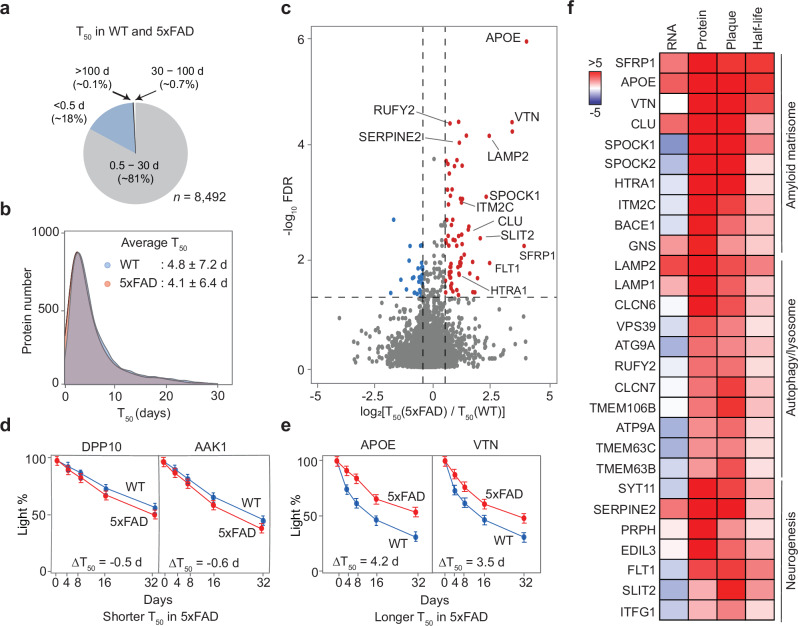
Murine models of Alzheimer's disease (AD) are crucial for elucidating disease mechanisms but have limitations in fully representing AD molecular complexities. Here we present the comprehensive, age-dependent brain proteome and phosphoproteome across multiple mouse models of amyloidosis. We identified shared pathways by integrating with human metadata and prioritized components by multi-omics analysis. Collectively, two commonly used models (5xFAD and APP-KI) replicate 30% of the human protein alterations; additional genetic incorporation of tau and splicing pathologies increases this similarity to 42%. We dissected the proteome-transcriptome inconsistency in AD and 5xFAD mouse brains, revealing that inconsistent proteins are enriched within amyloid plaque microenvironment (amyloidome). Our analysis of the 5xFAD proteome turnover demonstrates that amyloid formation delays the degradation of amyloidome components, including Aβ-binding proteins and autophagy/lysosomal proteins. Our proteomic strategy defines shared AD pathways, identifies potential targets, and underscores that protein turnover contributes to proteome-transcriptome discrepancies during AD progression.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures






Update of
-
Human-mouse proteomics reveals the shared pathways in Alzheimer's disease and delayed protein turnover in the amyloidome.bioRxiv [Preprint]. 2024 Oct 25:2024.10.25.620263. doi: 10.1101/2024.10.25.620263. bioRxiv. 2024. Update in: Nat Commun. 2025 Feb 11;16(1):1533. doi: 10.1038/s41467-025-56853-3. PMID: 39484428 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
