

The role of transposon activity in shaping cis-regulatory element evolution after whole-genome duplication
- PMID: 39939177
- PMCID: PMC11960703
- DOI: 10.1101/gr.278931.124
The role of transposon activity in shaping cis-regulatory element evolution after whole-genome duplication
Abstract
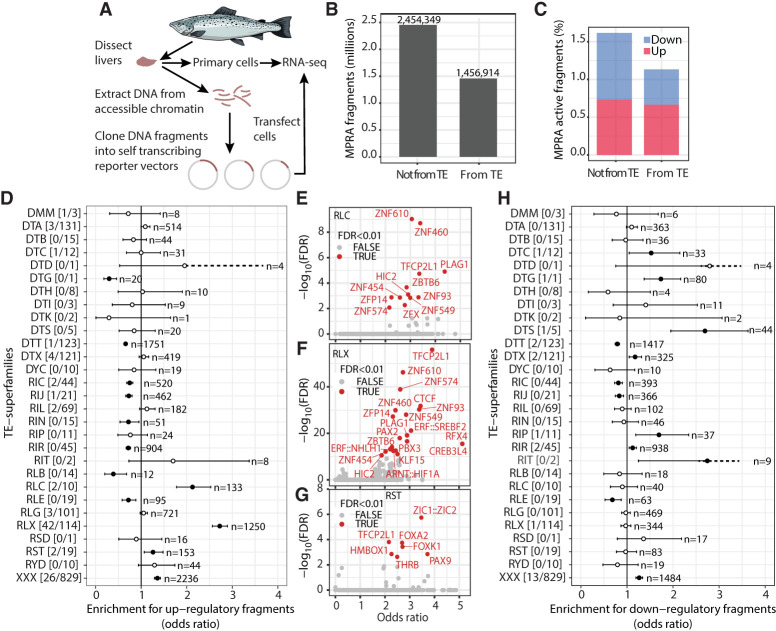
Whole-genome duplications (WGDs) and transposable element (TE) activity can act synergistically in genome evolution. WGDs can increase TE activity directly through cellular stress or indirectly by relaxing selection against TE insertions in functionally redundant, duplicated regions. Because TEs can function as, or evolve into, TE-derived cis-regulatory elements (TE-CREs), bursts of TE activity following WGD are therefore likely to impact evolution of gene regulation. Yet, the role of TEs in genome regulatory evolution after WGDs is not well understood. Here we used Atlantic salmon as a model system to explore how TE activity after the salmonid WGD ∼100 MYA shaped CRE evolution. We identified 55,080 putative TE-CREs using chromatin accessibility data from the liver and brain. Retroelements were both the dominant source of TE-CREs and had higher regulatory activity in MPRA experiments compared with DNA elements. A minority of TE subfamilies (16%) accounted for 46% of TE-CREs, but these "CRE superspreaders" were mostly active prior to the WGD. Analysis of individual TE insertions, however, revealed enrichment of TE-CREs originating from WGD-associated TE activity, particularly for the DTT (Tc1-Mariner) DNA elements. Furthermore, coexpression analyses supported the presence of TE-driven gene regulatory network evolution, including DTT elements active at the time of WGD. In conclusion, our study supports a scenario in which TE activity has been important in genome regulatory evolution, either through relaxed selective constraints or through strong selection to recalibrate optimal gene expression phenotypes, during a transient period following genome doubling.
© 2025 Monsen et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources