

Adenylate cyclase 10 promotes brown adipose tissue thermogenesis
- PMID: 39949963
- PMCID: PMC11821413
- DOI: 10.1016/j.isci.2025.111833
Adenylate cyclase 10 promotes brown adipose tissue thermogenesis
Abstract
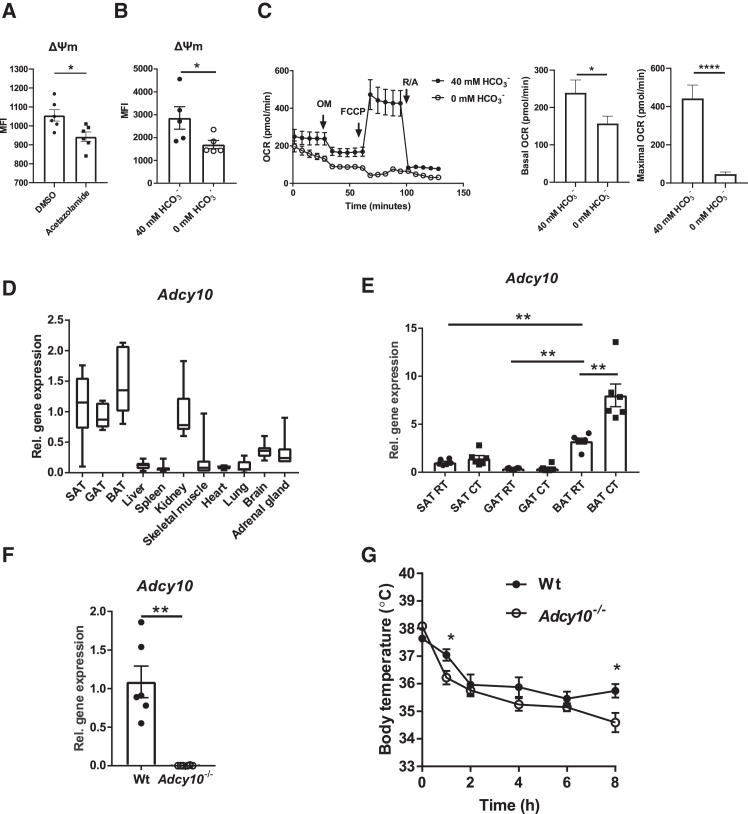
Brown adipose tissue (BAT) thermogenesis dissipates energy through heat production and thereby it opposes metabolic disease. It is mediated by mitochondrial membrane uncoupling, yet the mechanisms sustaining the mitochondrial membrane potential (ΔΨm) in brown adipocytes are poorly understood. Here we show that isocitrate dehydrogenase (IDH) activity and the expression of the soluble adenylate cyclase 10 (ADCY10), a CO2/bicarbonate sensor residing in mitochondria, are upregulated in BAT of cold-exposed mice. IDH inhibition or ADCY10 deficiency reduces cold resistance of mice. Mechanistically, IDH increases the ΔΨm in brown adipocytes via ADCY10. ADCY10 sustains complex I activity and the ΔΨm via exchange protein activated by cAMP1 (EPAC1). However, neither IDH nor ADCY10 inhibition affect uncoupling protein 1 (UCP1) expression. Hence, we suggest that ADCY10, acting as a CO2/bicarbonate sensor, mediates the effect of IDH on complex I activity through cAMP-EPAC1 signaling, thereby maintaining the ΔΨm and enabling thermogenesis in brown adipocytes.
Keywords: Cell biology; Molecular biology; Physiology.
© 2025 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
