

Creatine transporter (SLC6A8) knockout mice exhibit reduced muscle performance, disrupted mitochondrial Ca2+ homeostasis, and severe muscle atrophy
- PMID: 39952955
- PMCID: PMC11828924
- DOI: 10.1038/s41419-025-07381-x
Creatine transporter (SLC6A8) knockout mice exhibit reduced muscle performance, disrupted mitochondrial Ca2+ homeostasis, and severe muscle atrophy
Abstract
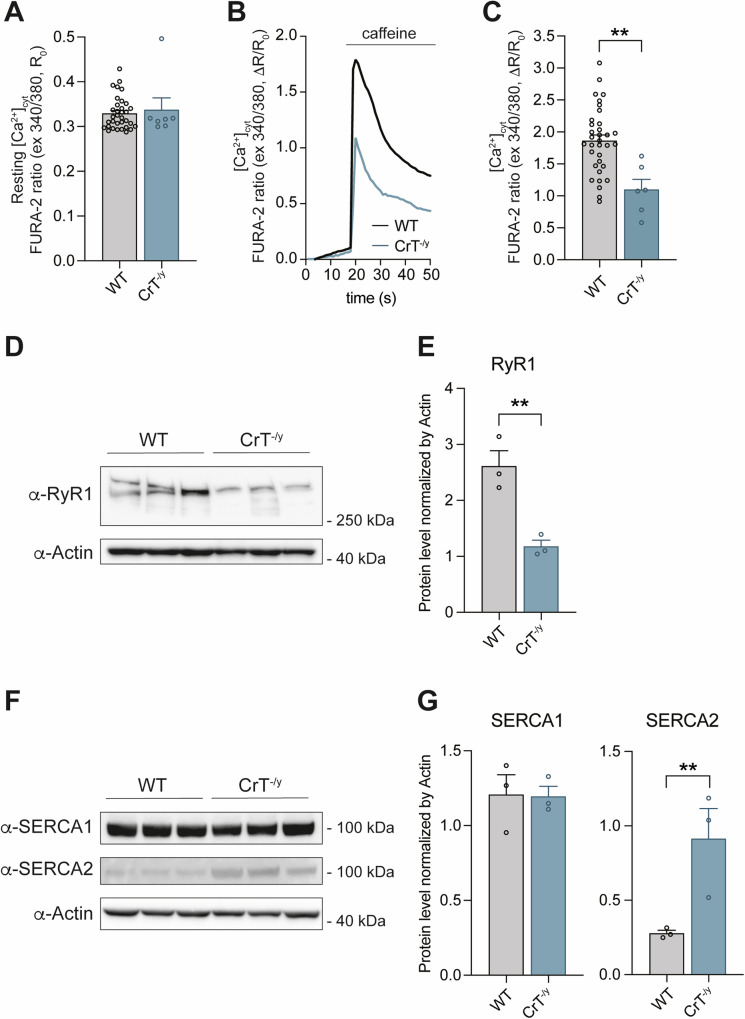
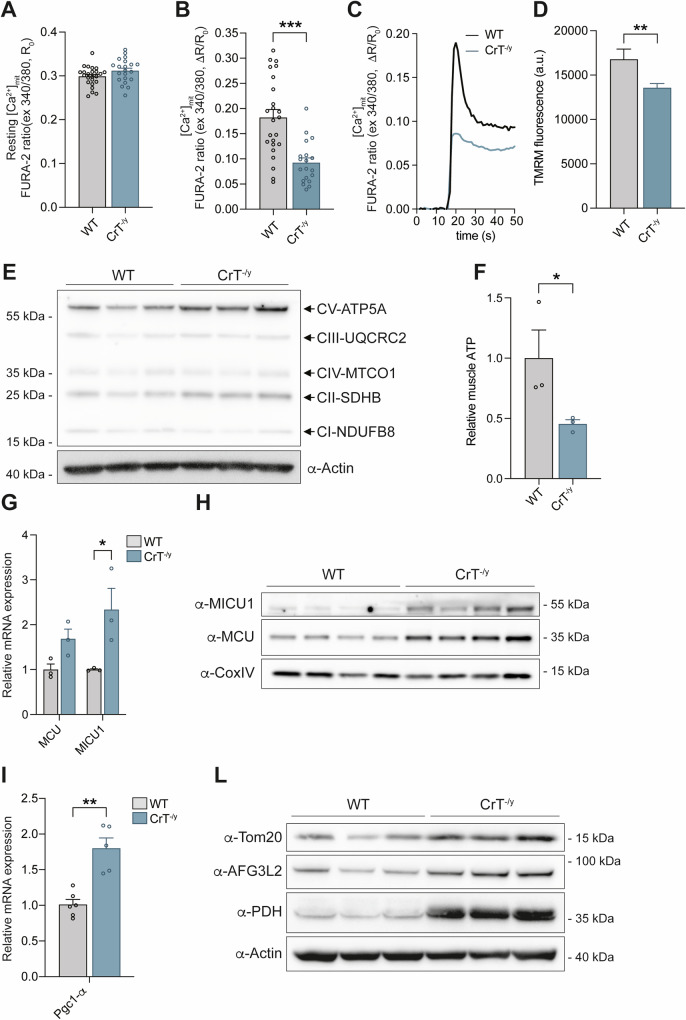
Creatine (Cr) is essential for cellular energy homeostasis, particularly in muscle and brain tissues. Creatine Transporter Deficiency (CTD), an X-linked disorder caused by mutations in the SLC6A8 gene, disrupts Cr transport, leading to intellectual disability, speech delay, autism, epilepsy, and various non-neurological symptoms. In addition to neurological alterations, Creatine Transporter knockout (CrT-/y) mice exhibit severe muscle atrophy and functional impairments. This study provides the first characterization of the skeletal muscle phenotype in CrT-/y mice, revealing profound ultrastructural abnormalities accompanied by reduced fiber cross-sectional area and muscle performance. Notably, mitochondria are involved, as evidenced by disrupted cristae, increased mitochondrial size, impaired Ca2+ uptake, reduced membrane potential and ATP production. Mechanistically, the expression of atrophy-specific E3 ubiquitin ligases and suppression of the IGF1-Akt/PKB pathway, regulated by mitochondrial Ca2+ levels, further support the atrophic phenotype. These findings highlight the profound impact of Cr deficiency on skeletal muscle, emphasizing the need for targeted therapeutic strategies to address both the neurological and peripheral manifestations of CTD. Understanding the underlying mechanisms, particularly mitochondrial dysfunction, could lead to novel interventions for this disorder.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval and consent to participate: All animal procedures were performed at the University of Padova and the University of Florence in accordance with the Italian law D. L.vo n.26/2014 and with the Directive 2010/63/EU of the European Parliament on the protection of animals used for scientific purposes. All methods were performed in accordance with the relevant guidelines and regulations.
Figures




References
-
- Wallimann T, Wyss M, Brdiczka D, Nicolay K, Eppenberger HM. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: the ‘phosphocreatine circuit’ for cellular energy homeostasis. Biochem J. 1992;281:21–40. - PMC - PubMed
-
- Ellington WR. Evolution and physiological roles of phosphagen systems. Annu Rev Physiol. 2001;63:289–325. - PubMed
-
- Brosnan ME, Edison EE, da Silva R, Brosnan JT. New insights into creatine function and synthesis. Adv Enzyme Regul. 2007;47:252–60. - PubMed
-
- Nash SR, Giros B, Kingsmore SF, Rochelle JM, Suter ST, Gregor P, et al. Cloning, pharmacological characterization, and genomic localization of the human creatine transporter. Recept Channels. 1994;2:165–74. - PubMed
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
