

Allosteric modulation and direct activation of glycine receptors by a tricyclic sulfonamide
- PMID: 39953280
- PMCID: PMC11828983
- DOI: 10.1038/s41598-025-90209-7
Allosteric modulation and direct activation of glycine receptors by a tricyclic sulfonamide
Abstract
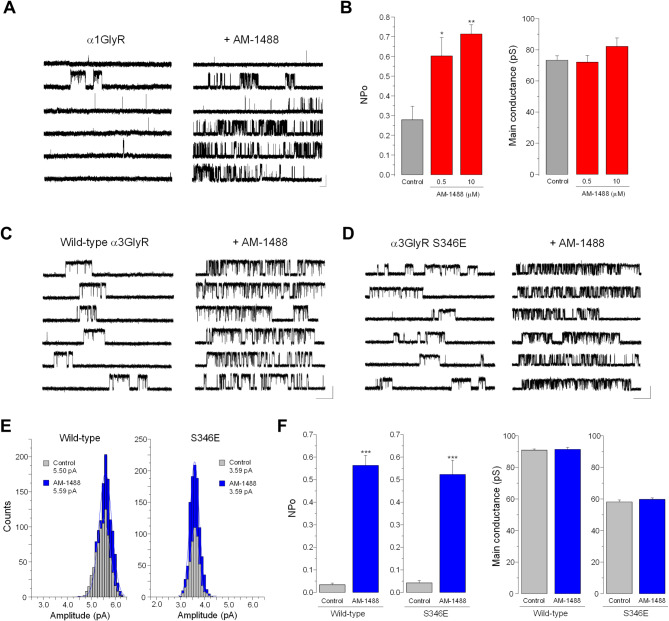
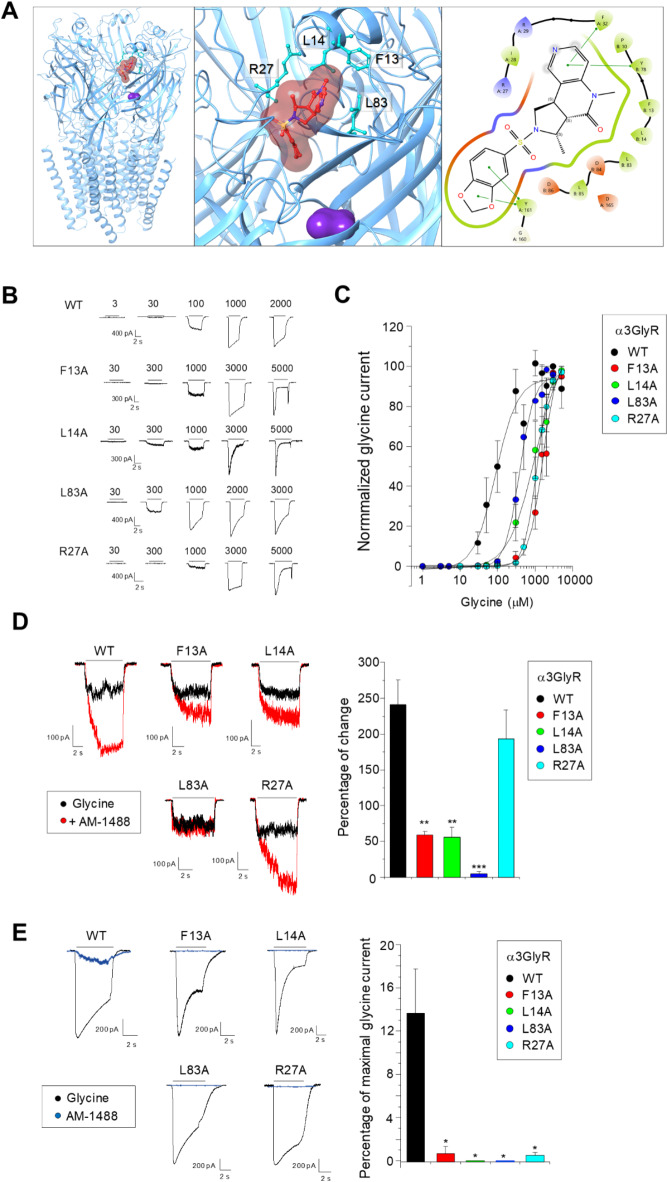
Ionotropic glycine receptors (GlyRs) are chloride-permeable ligand-gated ion channels expressed in the nervous system. Alterations to glycinergic inhibition and the generation of dysfunctional GlyRs have been linked to chronic pain, a widely prevalent disease. Positive allosteric modulators (PAMs) targeting GlyRs exerted analgesic effects, motivating research on glycinergic PAMs as potential pain therapies. Rationally designed tricyclic sulfonamides are novel glycinergic PAMs with analgesic activity. However, detailed electrophysiological studies on these PAMs are still limited, and the GlyR binding site structural data has not been yet validated by mutational studies. Here, we combined electrophysiology and bioinformatics to systematically study the AM-1488 actions, a prototypical tricyclic sulfonamide, on recombinant GlyRs. We determined that AM-1488 is a potent, non-selective PAM of mammalian GlyR subtypes. In addition, the compound displayed agonistic activity, with partial preference for α1GlyRs. Single channel assays revealed that the compound increased the channel open probability without changing conductance. Mutational analyses on the tricyclic sulfonamide site confirm the molecular determinants contributing to functional activity. Our findings further define the mechanistic framework underlying the GlyR modulation by this PAM class, suggesting that further structure-driven exploration within the tricyclic sulfonamide site may originate novel glycinergic modulators for future development.
Keywords: Allosteric modulation; Chronic pain; Electrophysiology; Glycine receptors; Molecular modelling; Pharmacology.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures


References
-
- Lynch, J. W. Native glycine receptor subtypes and their physiological roles. Neuropharmacology56(1), 303–309 (2009). - PubMed
-
- Nys, M., Kesters, D. & Ulens, C. Structural insights into cys-loop receptor function and ligand recognition. Biochem. Pharmacol.86(8), 1042–1053 (2013). - PubMed
-
- Lynch, J. W. et al. Glycine Receptor Drug Discovery Adv. Pharmacol., 79: 225–253. (2017). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
