

Anoctamin-1 is a core component of a mechanosensory anion channel complex in C. elegans
- PMID: 39956854
- PMCID: PMC11830769
- DOI: 10.1038/s41467-025-56938-z
Anoctamin-1 is a core component of a mechanosensory anion channel complex in C. elegans
Abstract
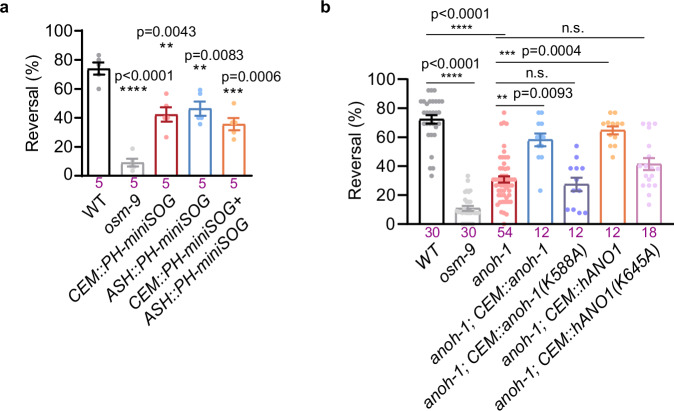
Mechanotransduction channels are widely expressed in both vertebrates and invertebrates, mediating various physiological processes such as touch, hearing and blood-pressure sensing. While previously known mechanotransduction channels in metazoans are primarily cation-selective, we identified Anoctamin-1 (ANOH-1), the C. elegans homolog of mammalian calcium-activated chloride channel ANO1/TMEM16A, as an essential component of a mechanosensory channel complex that contributes to the nose touch mechanosensation in C. elegans. Ectopic expression of either C. elegans or human Anoctamin-1 confers mechanosensitivity to touch-insensitive neurons, suggesting a cell-autonomous role of ANOH-1/ANO1 in mechanotransduction. Additionally, we demonstrated that the mechanosensory function of ANOH-1/ANO1 relies on CIB (calcium- and integrin- binding) proteins. Thus, our results reveal an evolutionarily conserved chloride channel involved in mechanosensory transduction in metazoans, highlighting the importance of anion channels in mechanosensory processes.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
- P40 OD010440/OD/NIH HHS/United States
- 31771113/National Natural Science Foundation of China (National Science Foundation of China)
- 31471023/National Natural Science Foundation of China (National Science Foundation of China)
- LZ22C090001/Natural Science Foundation of Zhejiang Province (Zhejiang Provincial Natural Science Foundation)
- 2021ZD0203303/Ministry of Science and Technology of the People's Republic of China (Chinese Ministry of Science and Technology)
LinkOut - more resources
Full Text Sources
Research Materials
