

Activated mTOR Signaling in the RPE Drives EMT, Autophagy, and Metabolic Disruption, Resulting in AMD-Like Pathology in Mice
- PMID: 39957408
- PMCID: PMC12151893
- DOI: 10.1111/acel.70018
Activated mTOR Signaling in the RPE Drives EMT, Autophagy, and Metabolic Disruption, Resulting in AMD-Like Pathology in Mice
Abstract
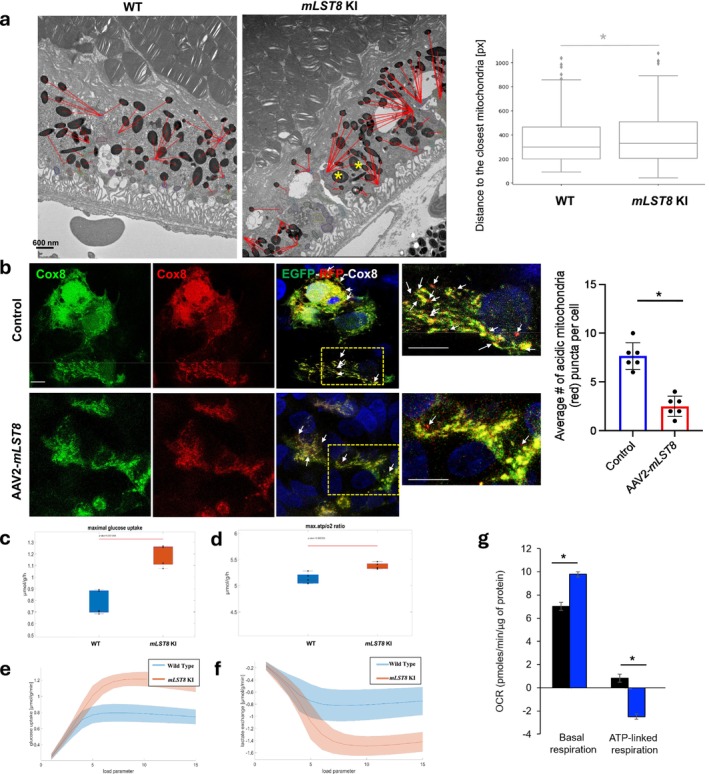
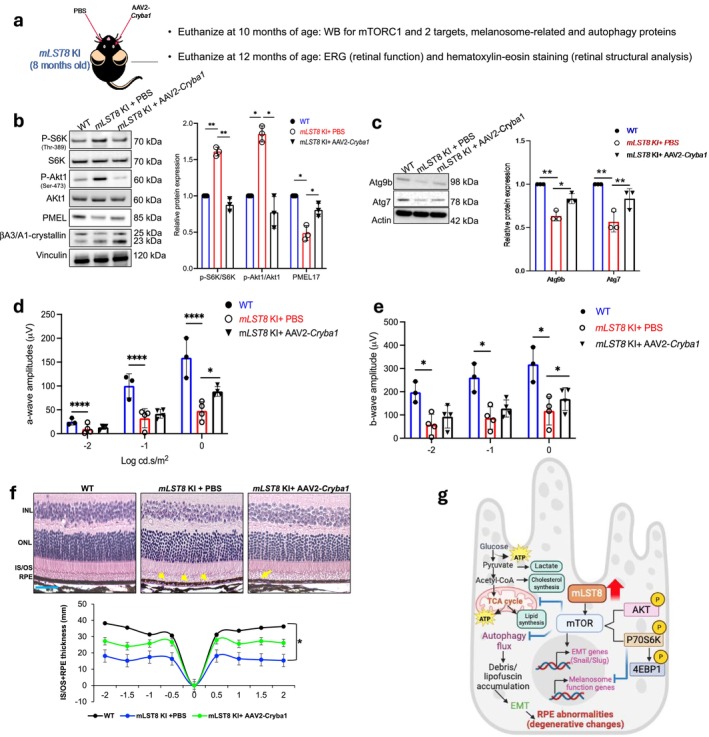
The mechanistic target of rapamycin (mTOR) complexes 1 and 2 (mTORC1/2) are crucial for various physiological functions. Although the role of mTORC1 in retinal pigmented epithelium (RPE) homeostasis and age-related macular degeneration (AMD) pathogenesis is established, the function of mTORC2 remains unclear. We investigated both complexes in RPE health and disease. Therefore, in this study, we have attempted to demonstrate that the specific overexpression of mammalian lethal with Sec13 protein 8 (mLST8) in the mouse RPE activates both mTORC1 and mTORC2, inducing epithelial-mesenchymal transition (EMT)-like changes and subretinal/RPE deposits resembling early AMD-like pathogenesis. Aging in these mice leads to RPE degeneration, causing retinal damage, impaired debris clearance, and metabolic and mitochondrial dysfunction. Inhibition of mTOR with TORIN1 in vitro or βA3/A1-crystallin in vivo normalized mTORC1/2 activity and restored function, revealing a novel role for the mTOR complexes in regulating RPE function, impacting retinal health and disease.
Keywords: RPE; epithelial–mesenchymal transition; mLST8; mTOR complex 1; mTOR complex 2; metabolic/mitochondrial changes.
© 2025 The Author(s). Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures




References
MeSH terms
Substances
Grants and funding
- University of Pittsburgh School of Medicine Jennifer Salvitti Davis, MD. Chair Professorship Start-up funding
- R01 EY031594/EY/NEI NIH HHS/United States
- R01EY031594/EY/NEI NIH HHS/United States
- K99 EY033421/EY/NEI NIH HHS/United States
- Research to Prevent Blindness Unrestricted funds to Pitt Dept of Ophthalmology
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
